

摘 要:【目的】比较研究塔里木河水系2种裂腹鱼类形态和骨骼系统解剖学的差异性,为高原鱼类的系统演化和地理分布格局提供科学参考。【方法】以新疆特有鱼类厚唇裂腹鱼和斑重唇鱼为研究对象,结合框架数据和传统可量数据进行形态特征和主成分分析;采用CT扫描法和煮沸剔肉法比较两者骨骼形态的差异。【结果】2种裂腹鱼31项性状中,1-2/体长(BL)差异显著(P lt; 0.05),体宽/体长(BW/BL)等25项比例性状差异极显著(P lt; 0.01);前2个主成分累计贡献率为56.74%,差异主要体现在机体头部和躯干前段。2种裂腹鱼的脑颅、咽颅和附肢骨骼数目相同,分别为42块、80块和132块;但两者的脊椎数和肋骨数存在一些不同:厚唇裂腹鱼脊柱4+42-43+1枚,肋骨20~21枚(2尾);斑重唇鱼脊柱4+34-35+1枚,肋骨17~18枚(2尾);尤其是第二前筛骨、蝶耳骨、翼耳骨、前颌骨、上颌骨、翼骨、中翼骨和咽骨上2种裂腹鱼的差异更明显。【结论】塔里木河水系2种裂腹鱼形态特征和骨骼形态差异较为明显,其差异和演化机制与青藏高原隆起有着密切联系,独特的形态进化特征和骨骼差异系统可作为鱼类分类鉴别的依据,丰富了其形态进化生物学资料。
关键词:塔里木河水系;厚唇裂腹鱼;斑重唇鱼;形态特征;骨骼解剖
中图分类号:S93 文献标志码:A 文章编号:1001-4330(2024)10-2566-10
收稿日期(Received):2024-04-06
基金项目:农业农村部财政专项(西北地区重点水域渔业资源与环境调查);新疆生产建设兵团重点领域科技攻关计划“南疆盐碱水渔业综合开发利用关键技术集成与示范”(2022DB019);塔里木畜牧科技兵团重点实验室项目“伊犁裂腹鱼人工繁殖关键技术研究”(HS201902)
作者简介:王程欣(1997-),女,山东日照人,硕士研究生,研究方向为鱼类生物学,(E-mail)m17862660810@163.com
通讯作者:陈生熬(1980-),男,青海西宁人,教授,博士,硕士生/博士生导师,研究方向为渔业资源与环境,(E-mail)chenshengao@163.com
0 引 言
【研究意义】鱼类外部形态特征差异是鉴别鱼类最基本方式,骨骼支撑和保护其机体并完成代谢功能[1-2],结构差异是适应不同环境的主要体现[3]。骨骼系统是基于外部形态结构的稳定性及在不同种类的不同特征,是鱼类形态及分类演化的重要依据[4-6]。裂腹鱼类是青藏高原三大自然鱼类类群组成之一,也是青藏高原隆起和地壳变化的参考依据。【前人研究进展】根据裂腹鱼栖息海拔高度的不同,将其分为原始、特化和高度特化三类[7-9]。世界上裂腹鱼类有 15 属 100 余种,中国约产 11 属 76 种(亚种)[10-11],我国分布于新疆塔里木河水系的厚唇裂腹鱼Schizothorax irregularis(Day 1876)和斑重唇鱼Diptychus maculatus(Steindachner 1866)均隶属裂腹鱼类,但由于地壳变化形成了不同属性的鱼类[12]。姚娜等[13]和张瑞等[14]研究斑重唇鱼和厚唇裂腹鱼的形态特征,武云飞[15]、赵海涛等[16]和王程欣等[17]对昆明裂腹鱼Schizothorax grahami、四川裂腹鱼S. kozlovi、塔里木裂腹鱼S. biddulphi和伊犁裂腹鱼S. pseudaksaiensis骨骼进行了系统解剖。【本研究切入点】目前针对裂腹鱼类研究较多,但对于骨骼解剖和系统分类等相结合阐述其演化的研究较少。特殊区域鱼类形态和解剖结构对于地壳演变和物种演化等有着重要的参考价值,故需关注鱼类形态和骨骼系统。【拟解决的关键问题】以新疆特有鱼类厚唇裂腹鱼和斑重唇鱼为研究对象,结合框架数据和传统可量数据的方法,分析形态特征;采用CT扫描法和煮沸剔肉法比较两者骨骼形态的差异,对其骨骼系统进行详细鉴别分析和比较,为高原鱼类的分类及系统进化研究提供理论依据。
1 材料与方法
1.1 材 料
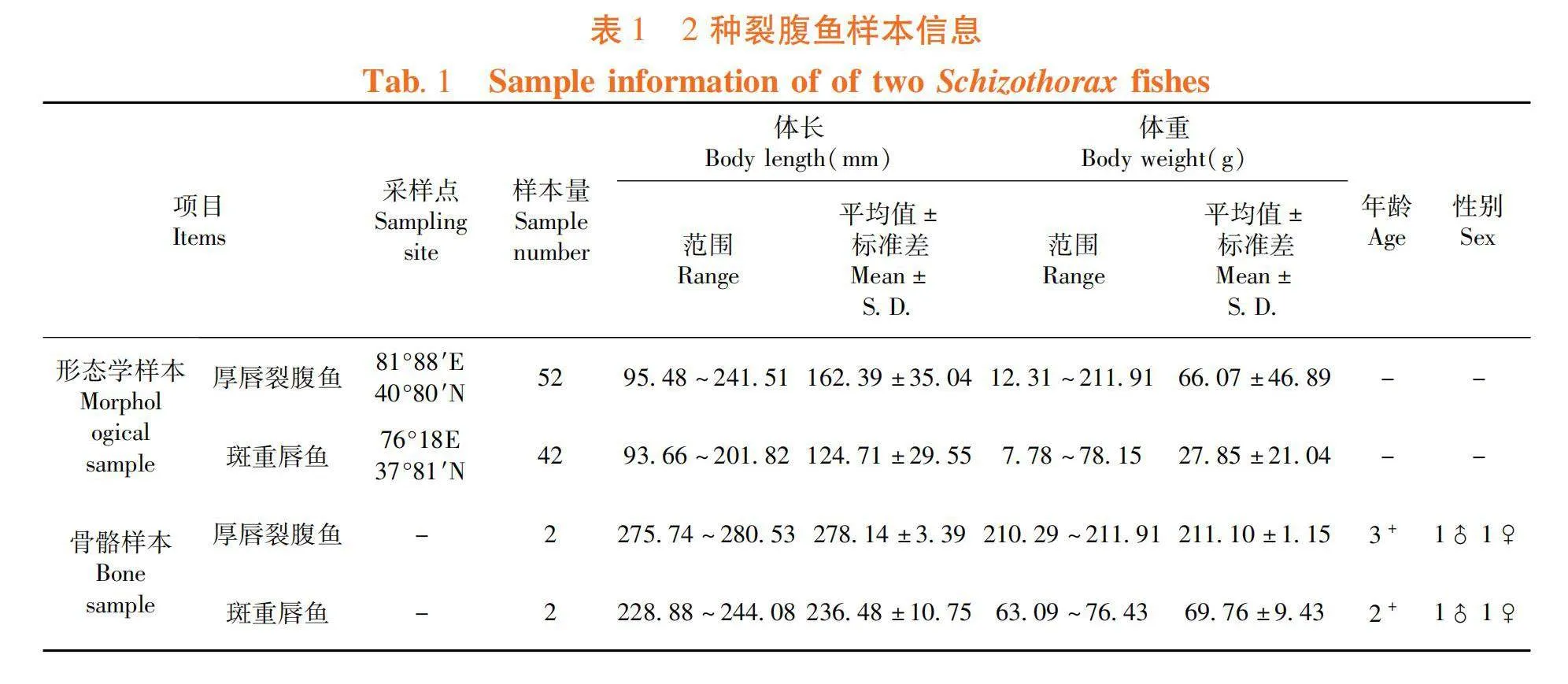
2022年5~7月,利用小抬网、地笼和定置刺网(网目 2 a = 2.00 cm)等工具,在新疆塔里木河水系采集厚唇裂腹鱼52尾和斑重唇鱼42尾;进行常规生物学测定后用福尔马林固定;挑选实验骨骼用鱼各2尾,均为体型外表完整的性成熟个体。表1
1.2 方 法
1.2.1 形态特征
使用传统形态学和框架法点对点测量鱼体形态特征,数显游标卡尺(CD67-S15PS)测量10项传统形态学数据和24个框架距离[18-19](精度0.01 mm),使用电子天平(LE403E)称量体质量(精度0.01 g)。图1
1.2.2 骨骼制备
CT扫描法:采用Micro CT μ80显微CT仪(型号)对全鱼样本进行360°旋转扫描拍照。
煮沸剔肉法:采用六步制作骨骼标本(煮肉、剔肉、浸蚀、脱脂、漂白和脱水等)。
1.3 数据处理
协方差矩阵提取主成分,得出贡献率,构建主成分散点图[20];SPSS 18.0 和 Origin 9.0软件分析数据;μCT Ray v4.0-1三维重建完整骨骼标本,解剖显微镜(SMZ-140 N2GG,MOTIC,厦门)观察并拍照记录(PowerShot A3000 IS,Canon,珠海)单个骨骼标本,Photoshop CS6处理后比较分析。
2 结果与分析
2.1 观察2种裂腹鱼外部形态特征并对比
研究表明,2种裂腹鱼体前部略呈圆筒形,后部侧扁;头圆锥形;吻钝,稍突出;下咽骨窄长,咽齿柱状,顶端尖且具钩。但厚唇裂腹鱼口形似马蹄,下颌无角质缘,下唇肥大,2对须;胸部鳞片埋于皮下,背鳍有硬刺,体无斑。斑重唇鱼口下位,横直或略弧形;下颌角质缘锐利;下唇表面粒状突起,左右两叶狭窄;1对须;胸、腹部裸露无鳞,背鳍无硬刺,奇鳍均具不规则褐色斑点。
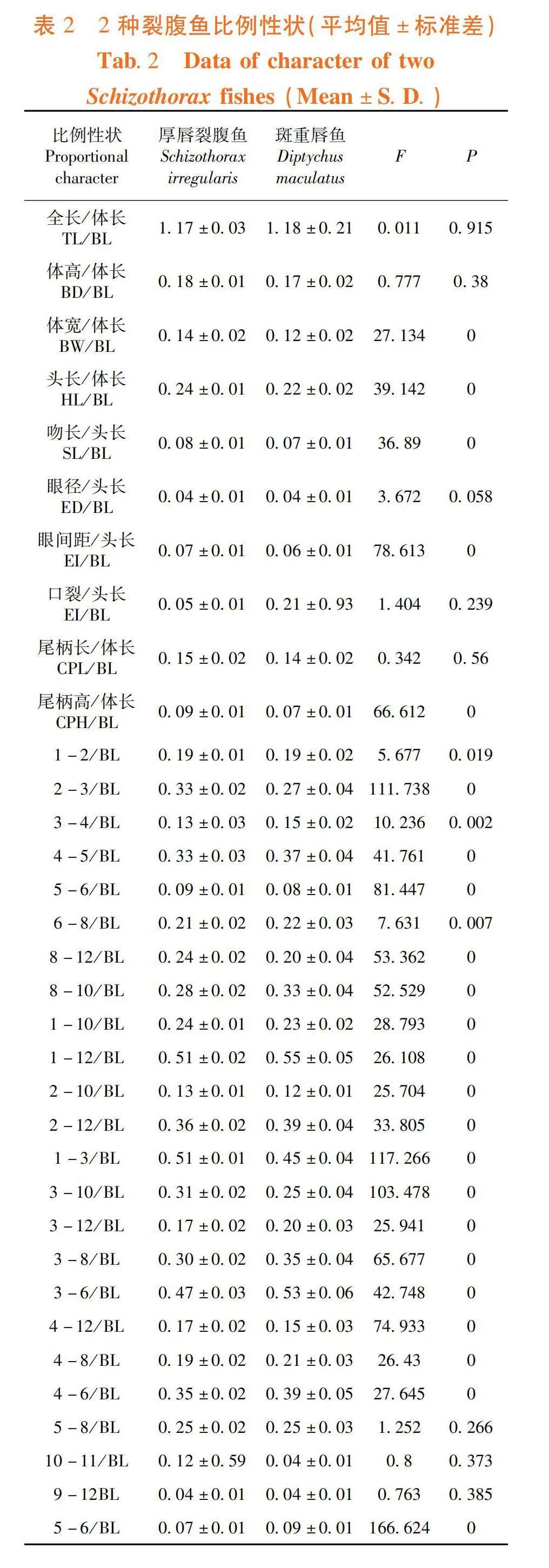
2.2 2种裂腹鱼传统形态度量性状
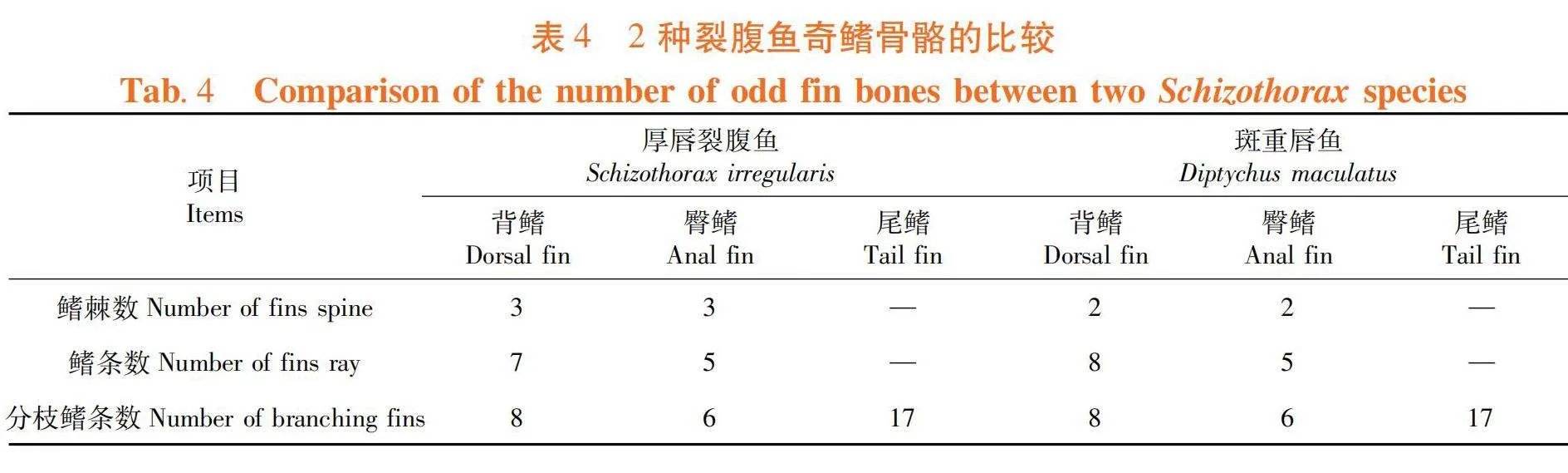
研究表明,1-2/BL在2种裂腹鱼间有显著差异(Plt;0.05),BW/BL等25项比例性状有极显著差异(P lt; 0.01)。表2,表3
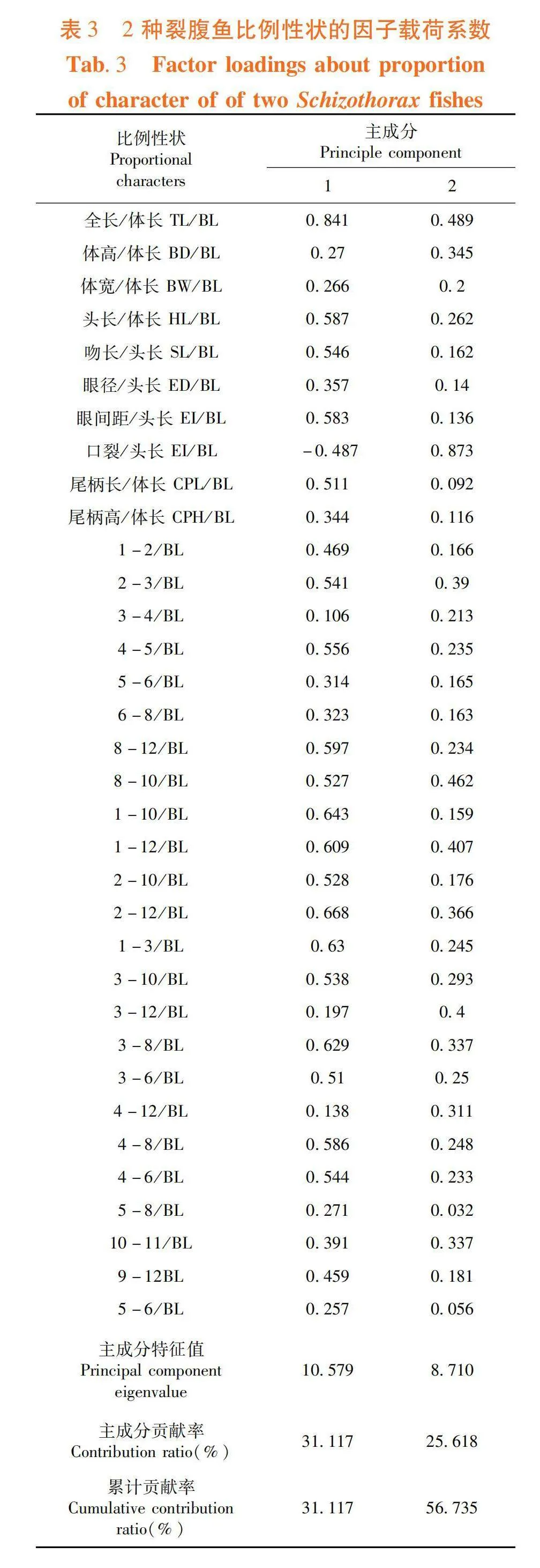
2.3 主成分贡献率
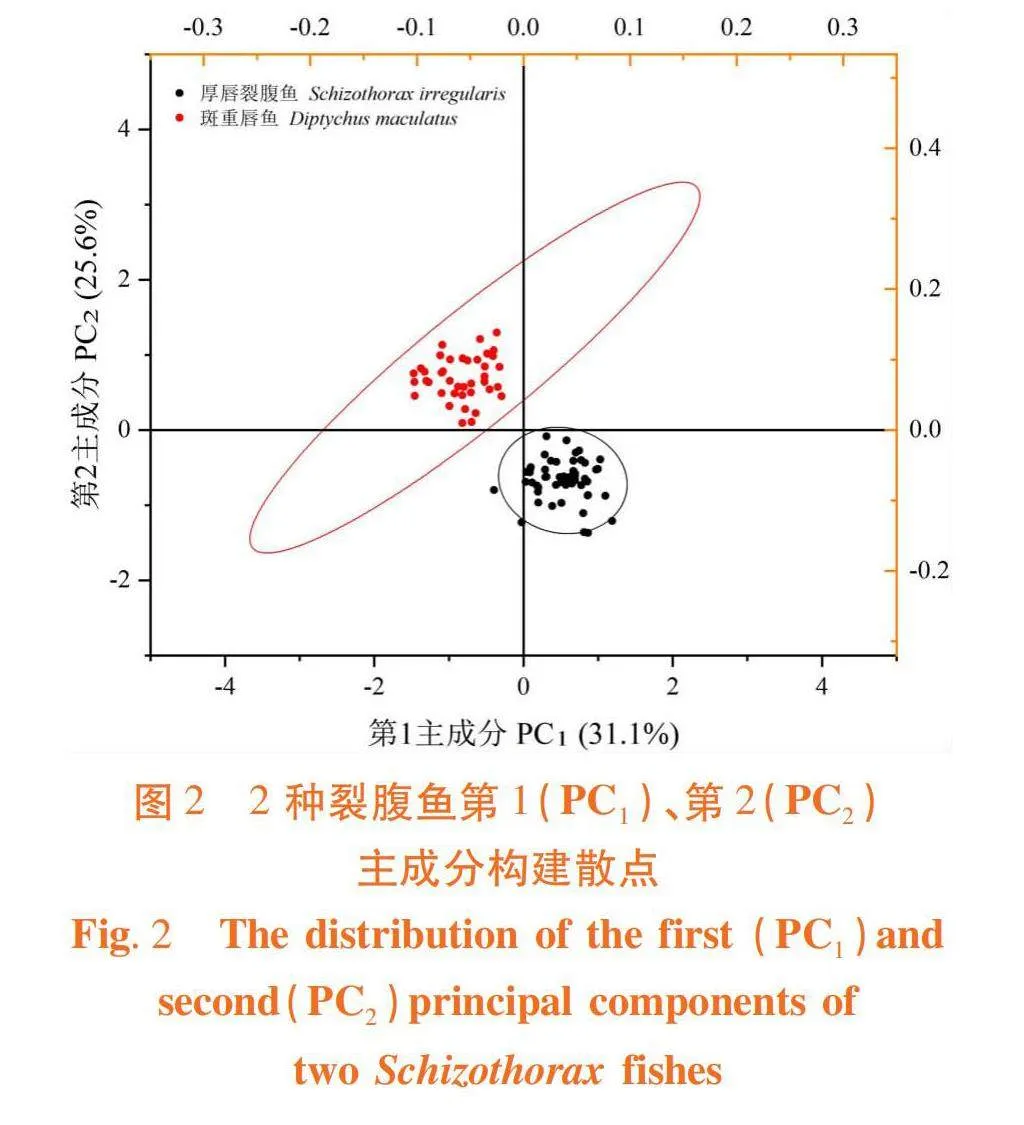
研究表明,2种裂腹鱼形态差异,前2个主成分达31.12%和25.62%的贡献率,累计贡献率达56.74%。主成分1载荷较大的参数有TL/BL、1-10/BL、1-12/BL、2-12/BL和3-8/BL,负荷值均高于0.600,主要反映纵向躯干部特征;主成分2中载荷较大的参数有EI/BL(gt; 0.600),主要反映头部和横向躯干部特征。
2种裂腹鱼间无重叠,虽有一定相似性,但存在明显差异。图2
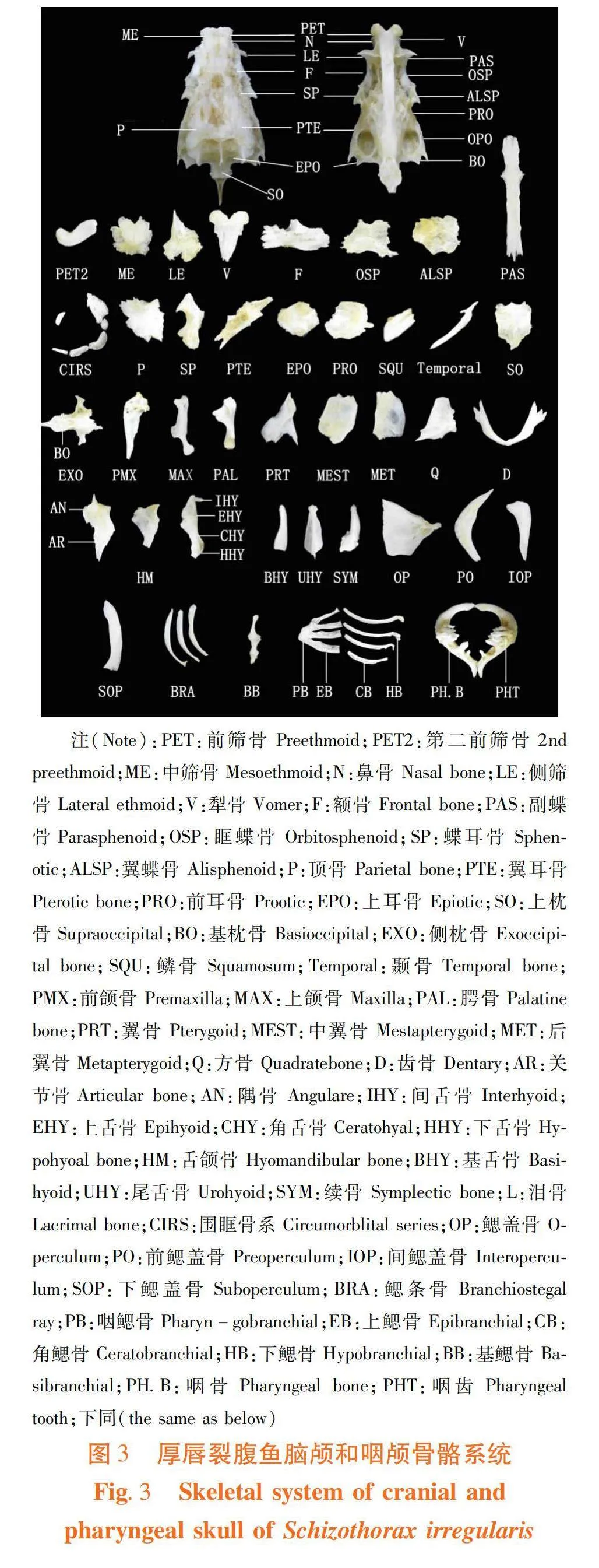
2.4 2种裂腹鱼脑颅解剖观察(图3)
2.4.1 鼻区
研究表明,裂腹鱼有9块骨组成。一对中筛骨基部两侧的鼻骨(N),一对头骨正前方的前筛骨(PET),介于前筛骨之间的第二前筛骨(PET2),脑颅背面前缘的中筛骨(ME),一对中筛骨两侧的侧筛骨(LE)和脑颅腹面最前方的犁骨(V)。
厚唇裂腹鱼第二前筛骨似“√”向内凹陷且弯曲度高,两端圆润突出;侧筛骨外侧缘下端中间部位形成明显凹陷。斑重唇鱼第二前筛骨似“棒”状,但一端向外延伸“尖”形凸出;侧筛骨外侧缘凹陷不显著。
2.4.2 眼区
研究表明,裂腹鱼有12块骨组成。一对脑颅背面的额骨(F),脑颅腹部中央的眶蝶骨(OSP),一对眼眶内后方的翼蝶骨(ALSP),脑颅底部中央的副蝶骨(PAS)和六块骨围成的围眶骨系(CIRS)。
厚唇裂腹鱼额骨前部1/2处骨片横向微凸,骨片较斑重唇鱼细长;翼蝶骨不规则状边缘凹陷明显;副蝶骨腹部前方“十”字形于骨块1/3处隆起,后部平直且长,左右对称;围眶骨系中泪骨较斑重唇鱼宽大。斑重唇鱼额骨呈“刀”形,骨片更宽;翼蝶骨边缘较曲折;副蝶骨腹部前方1/3处“十”字形部位较整体粗壮,后部2/3处又出现一明显凹陷,后“喇叭”状延伸,左右均等。
2.4.3 耳区
研究表明,裂腹鱼有14块骨组成。一对额骨后方的顶骨(P),一对眼眶后上方的蝶耳骨(SP),一对顶骨外部的翼耳骨(PTE),一对顶骨后方的上耳骨(EPO),一对颅腔侧面的前耳骨(PRO),一对于前耳骨、翼耳骨和侧枕骨间的后耳骨(OPO),一对脑后的鳞片骨(SQU)和一对连鳞片骨的颞骨(Temporale)。
厚唇裂腹鱼顶骨较宽,骨嵴延伸显著,两侧突出略呈对称;蝶耳骨一侧内凹明显呈“半圆”状,另一侧外突明显;翼耳骨一侧棱嵴凸起不明显,一端两后突大小相等;前耳骨较斑重唇鱼的有明显外突;鳞片骨整体扁平似“长方”形,边缘凹凸程度小;颞骨“尖刀”形骨片较斑重唇鱼的更延展细长。斑重唇鱼顶骨外边缘不规则凹陷较多,2种鱼顶骨中部有竖起骨嵴向四周延伸;蝶耳骨一侧上端略凹;翼耳骨一侧有2片且接连较紧的棱嵴凸起,两端向外呈“尖”状延伸;鳞片骨略细长,整体平直,边缘凹凸明显。
2.4.4 枕区
研究表明,裂腹鱼有6块骨组成。一对脑颅后中央的上枕骨(SO),一对基枕骨侧上方的侧枕骨(EXO),枕骨大孔(FOM)和颅骨最下方的基枕骨(BO)组成。
厚唇裂腹鱼上枕骨略宽,上枕嵴高耸,凹凸不明显略呈“”形。斑重唇鱼上枕骨边缘弯曲且外凸更明显略呈“△”状。
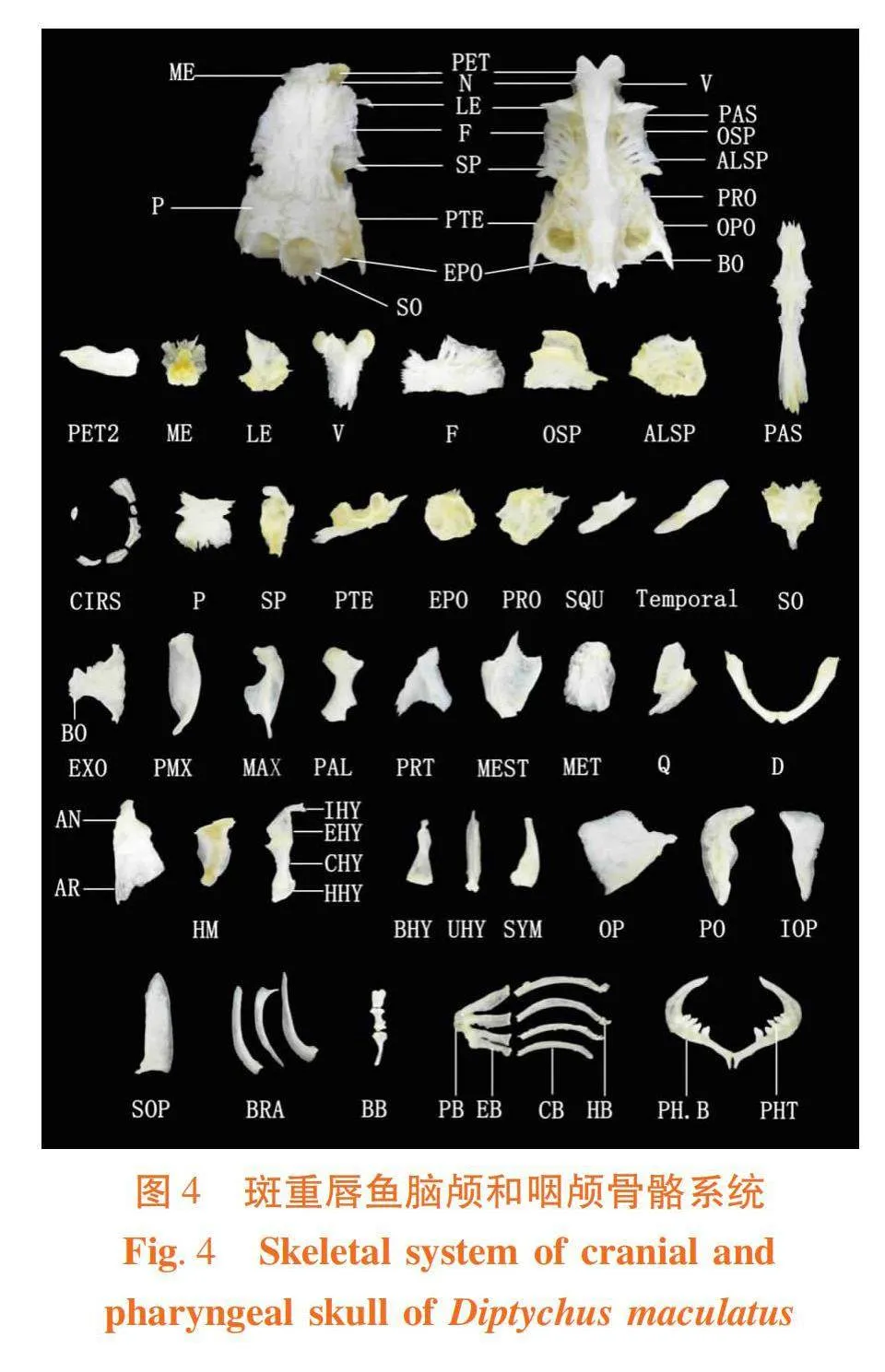
2.5 2种裂腹鱼咽颅解剖观察(图4~5)
2.5.1 颌弓区
研究表明,裂腹鱼有20块骨组成。一对位于吻前上方的前颌骨(PMX),一对前颌骨上方的上颌骨(MAX),一对犁骨两侧的腭骨(PAL),一对翼骨(PRT),一对翼骨下方的中翼骨(MEST),一对前鳃盖骨前方的后翼骨(MET),一对中翼骨腹缘的方骨(Q),一对下颌前部的齿骨(D),一对嵌在齿骨后端凹陷中的关节骨(AR)和一对与关节骨贴合的隅骨(AN)。
厚唇裂腹鱼前颌骨一端尖细无明显弯曲;上颌骨一侧呈方状凹陷,一端突出圆润呈柱状;腭骨纤细且棱角突出;翼骨外侧有尖锐突出;中翼骨呈方形。斑重唇鱼前颌骨一端有明显弯钩且弧度大;上颌骨一侧凹陷呈半圆状,一端有尖形突出;腭骨粗胖且钝;翼骨左右大致对称,外侧突出圆钝;中翼骨略呈“△”状,但一侧向外突出呈尖状。
2.5.2 舌弓区
研究表明,裂腹鱼有16块骨组成。一对鳃盖骨前上方的舌颌骨(HM)、一对上舌骨后上方的间舌骨(IHY)、一对角舌骨上缘的上舌骨(EHY)、一对上舌骨前方的角舌骨(CHY)、两对舌骨中部的下舌骨(HHY)、一块吻底部的基舌骨(BHY)、一块头骨下中央的尾舌骨(UHY)和一对方骨后方的续骨(SYM)。厚唇裂腹鱼角舌骨不及斑重唇鱼细长。
2.5.3 鳃盖骨区
研究表明,裂腹鱼有13块骨组成。一对头后部的鳃盖骨(OP)、一对前鳃盖骨(PO)、一对前鳃盖骨下部的间鳃盖骨(IOP)、鳃盖骨下部的下鳃盖骨(SOP)和三对鳃条骨(BRA)组成。
厚唇裂腹鱼前鳃盖骨似“镰刀”状,斑重唇鱼前鳃盖骨“镰刀”状凹陷处呈波浪形突起;厚唇裂腹鱼下鳃盖骨较斑重唇鱼更弯曲。
2.5.4 鳃弓区具四对,第五鳃弓变异成下咽骨。
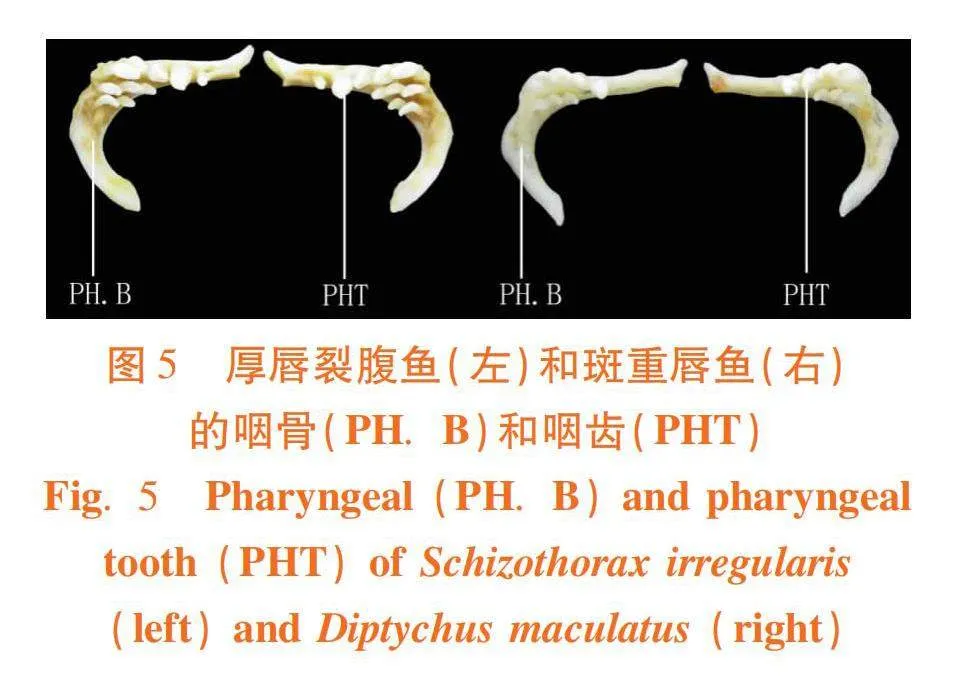
研究表明,裂腹鱼有31块骨组成。鳃弓中央的三块基鳃骨(BB)、基鳃骨两侧的三对下鳃骨(HB)、四对角鳃骨(CB)、四对连接在角鳃骨末端的上鳃骨(EB)、两对上鳃骨内侧的咽鳃骨(PB)和一对基鳃骨后端的咽骨(PH.B)。
厚唇裂腹鱼的齿式为2·3·5/5·3·2,其内侧有三排咽齿(PHT):第一排有最大的五枚,第二排的三枚和第三排的两枚较小;斑重唇鱼的齿式为3·4/4·3,其内侧有两排咽齿(PHT):第一排的四枚最大,第二排的三枚较小。2种裂腹鱼的咽骨和咽齿外形和数目有很大差异。
2.6 2种裂腹鱼躯干部和尾部解剖观察
2.6.1 脊柱和肋骨
研究表明,裂腹鱼前4枚愈合脊椎构成脊柱前部,呈平直板状且不发达的尾椎构成脊柱后部。厚唇裂腹鱼脊椎数为4+42-43+1,从第5椎体至第25或26椎体间有肋骨,数目为20或21;斑重唇鱼脊椎数为4+34-35+1,从第5椎体至第22或23椎体间有肋骨,数目为17或18。
2.6.2 附肢骨骼
研究表明,裂腹鱼奇鳍骨骼均不成对。背鳍和臀鳍,两者仅由支鳍骨支持;尾鳍因有脊柱末端等的加入使其骨骼更为复杂。表4
偶鳍骨骼均成对,胸鳍由1鳍棘,15鳍条组成;腹鳍由1鳍棘,8鳍条组成。胸鳍鳍条基部连肩带,肩带含肩胛骨(1对)、匙骨(1对)、上匙骨(1对)、后匙骨(1对)、乌喙骨(1对)和中乌喙骨(1对)。四对胸鳍支鳍骨分别与肩胛骨和乌喙骨相连,后接鳍条;腹鳍腰带左右成对,其支鳍骨后缘与鳍条相连。表5
3 讨 论
3.1 2种裂腹鱼外部形态特征差异
鱼体形态特征受遗传与环境双重因素影响,是鉴别分类的重要依据[21]。厚唇裂腹鱼和斑重唇鱼外部形态较为相似,但斑重唇鱼触须明显退化为1对,可能与鱼类栖息的海拔高度及摄食行为不同密切相关。需在较短的摄食期内获得充足食物供给,扩大摄食种类,增大摄食量,触须退化,感知功能减弱以增大其食谱范围[22]。
裂腹鱼进化适应,并随高原上升演变为原始或特化等级,而斑重唇鱼则演化到特化等级,触须数目发生变化[22]。此外,斑重唇鱼栖息海拔高于厚唇裂腹鱼,与特化等级一致[12]。2种裂腹鱼从原始等级到特化等级的演化过程中,经历过高原海拔急剧上升和栖息环境变化过程,而出现不同形态特征异同。
3.2 2种裂腹鱼头部骨骼差异
脊椎动物中鱼类具有最复杂的骨骼系统[23],是系统分类学研究重要依据。鱼类头部的形态构造反映其在生态系统中定位,同时其骨骼构造反映其进化过程[24-25]。头部骨骼包括保护脑的脑颅和支持口器、咽部的咽颅两部分[26]。厚唇裂腹鱼、斑重唇鱼和叶尔羌高原鳅(Triplophysa yarkandensis)[27]都分布于塔里木河水系,厚唇裂腹鱼和斑重唇鱼脑颅的骨骼数目都是42块,侧枕骨位于基枕骨侧上方,鳞骨位于脑后部,翼蝶骨外接蝶耳骨;而后者脑颅骨骼数目为25块,且无侧枕骨,鳞骨退化,翼蝶骨外接翼耳骨,同一流域不同属间骨骼数目和位置存在差异。不同流域同属间骨骼的形态也存在不同,研究中厚唇裂腹鱼蝶耳骨一侧明显内凹成“半圆”状,另一侧外突明显,鳞片骨扁宽似“长方”形,边缘凹凸程度小,不同于对伊犁河水系伊犁裂腹鱼的骨骼解剖研究[17],该河段伊犁裂腹鱼蝶耳骨外侧缘较圆滑,外突不发达,鳞片骨相较而言略细长。
咽颅作为摄食器官重要组成部分,形态变化受摄食习性和栖息环境等影响更大[28]。研究发现,同为刮食性的裂腹鱼类,厚唇裂腹鱼和塔里木裂腹鱼翼骨底部凹陷处偏向一侧的1/3处,而斑重唇鱼和伊犁裂腹鱼翼骨大致对称。虽有相似咽骨和咽齿外形,但塔里木裂腹鱼、伊犁裂腹鱼、四川裂腹鱼和昆明裂腹鱼的咽骨两端更尖锐,这与它们杂食性偏肉类型的摄食习性有关。此外,随鱼类摄食种类不同,下咽齿的形状和功能相应有所差异,厚唇裂腹鱼下咽齿为3行,斑重唇鱼为2行;斑重唇鱼下咽齿行数减少与其杂食性程度更宽有关;鱼类外侧一行咽齿在摄食中起主要作用,但随杂食程度的加强,最内侧一行下咽齿(仅2枚)研磨食物功能显著降低,甚至退化消失。因此,下咽齿行数在其特定栖息环境下发生的变异进一步验证了2种裂腹鱼进化和演变。
3.3 2种裂腹鱼脊椎骨及附肢骨骼差异
脊柱是鱼类机体主要的支撑骨骼,不同鱼类脊柱差异较大[29]。鲤科鱼类的生态习性及体型与脊椎骨数明显相关[30],鱼类从初孵仔鱼至成鱼的过程,遗传和环境等均可影响其骨骼发育[31]。厚唇裂腹鱼和斑重唇鱼同为裂腹鱼亚科鱼类,但脊椎骨和肋骨数具显著差异(P lt; 0.05),其脊椎骨和肋骨与其他裂腹鱼[16-17]的比较。厚唇裂腹鱼多生活于水流较急的叶尔羌河中游段深水河床中,斑重唇鱼栖息于叶尔羌河段的上游高海拔水流较缓,如回水湾或深水池潭等[12],不同生态位致使其适应不同而变化[32]。
鱼类尾部骨骼发育提供了不同发育时期对功能趋势和环境偏好的见解[33]。厚唇裂腹鱼在湍急水流中快速游泳捕获猎物,因其相对偏少的肋骨数和较多的尾椎数,加之发达的尾部,为捕食爆发性的游动提供骨骼结构支持[29]。斑重唇鱼隶属重唇鱼属且喜生活于水流较缓处[12],是造成脊椎骨和肋骨数明显少于厚唇裂腹鱼的原因。塔里木裂腹鱼和伊犁裂腹鱼喜栖息于静水河道[12,15],肋骨数较多于厚唇裂腹鱼;而四川裂腹鱼和昆明裂腹鱼生活于金沙江等流域[34],相对更多的脊椎骨和肋骨数可看出四川裂腹鱼和昆明裂腹鱼生活的金沙江水流较塔里木河更为湍急,也在有关研究中得到验证[35-36]。
4 结 论
2种裂腹鱼类外部形态差异明显,尤其体现在吻部和触须上。两者的31项生物学性状比例中,25项比例性状存在显著差异(P lt; 0.01),其差异主要体现在机体头部和躯干前段。2种裂腹鱼全身骨骼系统的差异较小;两者的脑颅、咽颅和附肢骨骼数目相同,但脊椎数和肋骨数存在一些不同,尤其在第二前筛骨、蝶耳骨、翼耳骨、前颌骨、上颌骨、翼骨、中翼骨和咽骨上差异更明显。2种裂腹鱼类独特的形态进化特征和骨骼差异与青藏高原的隆起有着密切联系。
参考文献(References)
[1]李仲辉, 马云霞, 杨太有. 长尾大眼鲷骨骼学的研究[J]. 水产科学, 2012, 31(12): 741-744.
LI Zhonghui, MA Yunxia, YANG Taiyou. Observation of skeleton system in spotfin bigeyePriacanthus tayenusRichardson[J]. Fisheries Science, 2012, 31(12): 741-744.
[2] 刘钊, 赵仲孟, 杨世勇, 等. 西伯利亚鲟幼鱼骨骼系统解剖研究[J]. 水生生物学报, 2019, 43(1): 117-122.
LIU Zhao, ZHAO Zhongmeng, YANG Shiyong, et al. The anatomy of the skeletal system of acipenser baeri juvenile[J]. Acta Hydrobiologica Sinica, 2019, 43(1): 117-122.
[3]Whitney J L, Donahue M J, Karl S A. Niche divergence along a fine-scale ecological gradient in sympatric color morphs of a coral reef fish[J]. Ecosphere, 2018, 9(1): e02015.
[4] 孟庆闻. 鱼类比较解剖[M]. 北京: 科学出版社, 1987: 57-118.
MENG Qingwen. Comparative Anatomy of fish[M]. Beijing: Science Press, 1987: 57-118.
[5] 宋小广, 刘海平, 刘艳超, 等. 双须叶须鱼脑颅骨骼形态学的研究[J]. 高原农业, 2019, 3(2): 159-167.
SONG Xiaoguang, LIU Haiping, LIU Yanchao, et al. Morphology of neurocranium skeleton in Ptychobarbus dipogon[J]. Journal of Plateau Agriculture, 2019, 3(2): 159-167.
[6] 吕雪娇, 王雨浓, 刘清华, 等. 鞍带石斑鱼仔稚幼鱼骨骼发育与生长特性研究[J]. 海洋科学, 2018, 42(5): 116-121.
LYU Xuejiao, WANG Yunong, LIU Qinghua, et al. Research on skeletal development and allometric growth in larval and juvenile Epinephelus lanceolatus[J]. Marine Sciences, 2018, 42(5): 116-121.
[7] 马宝珊, 谢从新, 霍斌, 等. 裂腹鱼类生物学研究进展[J]. 江西水产科技, 2011, (4): 36-40.
MA Baoshan, XIE Congxin, HUO Bin, et al. Research Progress in Biology of Schizothorax Fishes[J]. Jiangxi Fishery Science and Technology, 2011, (4): 36-40.
[8] 云飞. 中国裂腹鱼亚科鱼类的系统分类研究[J]. 高原生物学集刊, 1984, (12): 120-140.
WU Yunfei. A Systematic Taxonomic Study of Schizothoracinae in China[J]. Journal of Plateau Biology, 1984, (12): 120-140.
[9] 毅峰, 曹文宣. 裂腹鱼亚科鱼类[M]. 北京: 科学出版社, 2000.
CHEN Yifeng, CAO Wenxuan. Fishes of Schizothoracinae[M]. Beijing: Science Press, 2000.
[10] 陈娟. 青藏高原裂腹鱼亚科鱼类系统发育关系研究[D]. 北京: 中国科学院大学, 2016.
CHEN Juan. Phylogenetic Relationship of Schizothoracinae in Tibetan Plateau[D]. Beijing: University of Chinese Academy of Sciences, 2016.
[11] Mirza M R. A contribution to the systematics of the Schizothoracine fishes (Pisces: Cyprinidae) with the description of three new tribes[J]. Pakistan Journal of Zoology, 1991, 23: 339-341.
[12] 郭焱. 新疆鱼类志[M]. 乌鲁木齐: 新疆科学技术出版社, 2012.
GUO Yan. Xinjiang of Fishery[M]. Urumqi: Xinjiang Science and Technology Press, 2012.
[13] 姚娜, 葛建民, 王程欣, 等. 特克斯河斑重唇鱼生态学特征[J]. 新疆农业科学, 2023, 60(7): 1798-1805.
YAO Na, GE Jianmin, WANG Chengxin, et al. Ecological characteristics of Diptychus maculates in Turks River[J]. Xinjiang Agricultural Sciences, 2023, 60(7): 1798-1805.
[14] 张瑞, 赵贺, 聂竹兰, 等. 新疆克孜勒河4种裂腹鱼形态差异比较[J/OL]. 水生态学杂志: 1-12.
ZHANG Rui, ZHAO He, NIE Zhulan, et al. Comparison of morphological differences among four species of Schizothorax in the Kizil River, Xinjiang[J/OL]. Journal of Hydroecology: 1-12.
[15] 武云飞, 吴翠珍. 青藏高原鱼类[M]. 成都: 四川科学技术出版社, 1982.
WU Yunfei, WU Cuizhen. The fishes of the Qinghai-Xizang Plateau[M]. Chengdu: Sichuan Scientific amp; Technical Publishers, 1982.
[16] 赵海涛, 陈永祥, 胡思玉, 等. 四川裂腹鱼与昆明裂腹鱼骨骼系统的比较研究[J]. 安徽农业科学, 2011, 39(22): 13848-13853, 13873.
ZHAO Haitao, CHEN Yongxiang, HU Siyu, et al. Comparative study on the skeletal system of Schizothorax kozlovi and Schizothorax grahami[J]. Journal of Anhui Agricultural Sciences, 2011, 39(22): 13848-13853, 13873.
[17] 王程欣, 葛建民, 王新月, 等. 2种裂腹鱼类骨骼结构比较研究[J]. 四川动物, 2023, 42(4): 430-438.
WANG Chengxin, GE Jianmin, WANG Xinyue, et al. Comparative study on skeletal structure of two species of schizothoracinae[J]. Sichuan Journal of Zoology, 2023, 42(4): 430-438.
[18] 李思发, 李晨虹, 李家乐. 尼罗罗非鱼品系间形态差异分析[J]. 动物学报, 1998, 44(4): 450-457.
LI Sifa, LI Chenhong, LI Jiale. Analysis of morphological variations among strains of Nile tilapia (oreochromis niloticus)[J]. Current Zoology, 1998, 44(4): 450-457.
[19] Bookstein F L. Size and shape spaces for landmark data in two dimensions]: rejoinder[J]. Statistical Science, 1986, 1(2): 238-242.
[20] 李振通, 成美玲, 田永胜, 等. 杂交种“云龙石斑鱼” 与亲本的形态差异分析[J]. 渔业科学进展, 2019, 40(4): 73-83.
LI Zhentong, CHENG Meiling, TIAN Yongsheng, et al. Analysis of the Morphological Differences Between Hybrid “Epinephelus moara♀ × E. lanceolatus♂” and Its Parents[J]. Progress in Fishery Sciences, 2019, 40(4): 73-83.
[21] Kinsey S T, Orsoy T, Bert T M, et al. Population structure of the Spanish sardine Sardinella aurita: natural morphological variation in a genetically homogeneous population[J]. Marine Biology, 1994, 118(2): 309-317.
[22] 中国科学院青藏高原综合科学考察队. 青藏高原隆起的时代、幅度和形式问题[M]. 北京: 科学出版社, 1981.
Chinese Academy of Sciences Tibetan Plateau comprehensive scientific expedition team. The Era, amplitude and Form of the Tibetan Plateau Uplift[M]. Beijing: Science Press, 1981.
[23] 黄永彬, 张财文, 焦润杰, 等. 大圆颚针鱼骨骼标本的制作[J]. 生物学教学, 2019, 44(1): 35-36.
HUANG Yongbin, ZHANG Caiwen, JIAO Runjie, et al. Preparation of the skeletal specimen of Tylosurus giganteus[J]. Biology Teaching, 2019, 44(1): 35-36.
[24] 李仲辉, 靳萍, 杨太有. 斑鳍方头鱼骨骼学研究[J]. 河南师范大学学报(自然科学版), 2012, 40(2): 137-139.
LI Zhonghui, JIN Ping, YANG Taiyou. Skeleton system ofBranchiostegus auratus[J]. Journal of Henan Normal University (Natural Science Edition), 2012, 40(2): 137-139.
[25] 杨太有, 李仲辉. 二长棘鲷和黄鳍鲷骨骼系统的比较[J]. 广东海洋大学学报, 2008, 28(3): 1-5.
YANG Taiyou, LI Zhonghui. A study on osteology of Parargyrops edita Tanaka and Sparus latus houttuyn[J]. Journal of Guangdong Ocean University, 2008, 28(3): 1-5.
[26] 谢从新. 鱼类学[M]. 北京: 中国农业出版社, 2010.
XIE Congxin. Ichthyology[M]. Beijing: China Agriculture Press, 2010.
[27] 陈生熬, 谢从新, 王智超, 等. 叶尔羌高原鳅脑颅解剖鉴定分析[J]. 渔业科学进展, 2014, 35(1): 41-45.
CHEN Shengao, XIE Congxin, WANG Zhichao, et al. The skull anatomy of Triplophysa yarkandensis (Day)[J]. Progress in Fishery Sciences, 2014, 35(1): 41-45.
[28] 殷名称. 鱼类生态学[M]. 北京: 中国农业出版社, 1995.
YIN Mingcheng. Fish Ecology[M]. Beijing: China Agriculture Press, 1995.
[29] 杨蕊, 于刚, 胡静, 等. 青干金枪鱼骨骼系统研究[J]. 南方水产科学, 2021, 17(2): 36-43.
YANG Rui, YU Gang, HU Jing, et al. Research on skeleton system ofThunnus tonggol[J]. South China Fisheries Science, 2021, 17(2): 36-43.
[30] 王永梅, 唐文乔. 中国鲤形目鱼类的脊椎骨数及其生态适应性[J]. 动物学杂志, 2014, 49(1): 1-12.
WANG Yongmei, TANG Wenqiao. The number of vertebrae from Chinese species of the Cypriniformes and its ecological adaptation[J]. Chinese Journal of Zoology, 2014, 49(1): 1-12.
[31] Cahu C, Zambonino Infante J, Takeuchi T. Nutritional components affecting skeletal development in fish larvae[J]. Aquaculture, 2003, 227(1/2/3/4): 245-258.
[32] 曹晓颖. 鳜鱼骨骼结构、骨化发育及颌骨重塑的初步研究[D]. 上海: 上海海洋大学, 2019.
CAO Xiaoying. A Preliminary Study on the Skeletal Structure, ossification Development and Jaw Remodeling of Mandarin Fish[D]. Shanghai: Shanghai Ocean University, 2019.
[33] Fukuhara A O. Study on the development of functional morphology and behaviour of the larvae of eight commercially valuable teleost fishes[J]. Contrib Fish Res Jpn Sea Block, 1992, 25: 1-22.
[34] 伍献文. 中国鲤科鱼类志(下卷)[M]. 上海: 上海科学技术出版社, 1995: 65-66.
WU Xianwen. Annals of Cyprinid Fishes of China (Part II) [M]. Shanghai: Shanghai Science and Technology Press, 1995: 65-66.
[35] 茹辉军, 张燕, 吴湘香, 等. 金沙江下游沿岸区大型底栖动物群落结构及影响因素[J]. 中国水产科学, 2022, 29(12): 1679-1692.
RU Huijun, ZHANG Yan, WU Xiangxiang, et al. Macrozoobenthos community structure and its relationship with environmental factors in the riparian zone of the lower Jinsha River[J]. Journal of Fishery Sciences of China, 2022, 29(12): 1679-1692.
[36] 艾尼瓦尔·牙生. 塔里木河干流标准水量监测断面试点建设分析[J]. 陕西水利, 2020, (1): 52-53.
Ainivaer Yasheng. Analysis on pilot construction of standard water quantity monitoring section of Tarim River[J]. Shaanxi Water Resources, 2020, (1): 52-53.
Comparative study on morphological and skeletal differences of two Schizothorax species in Tarim River system
WANG Chengxin1,HU Linghui1,YANG Liting1,LIN Xuyuan2,HU Bolin2,
YUAN Zhichang3,SONG Yong1,CHEN Shengao1
(1. Tarim Rare Fish Research Center/College of Life Science and Technology/Tarim University, Aral Xinjiang 843300, China; 2 Aquatic Technology Extension Station of Xinjiang Production and Construction Corps, Urumqi 830000, China; 3. College of Fisheries and Life Sciences, Shanghai Ocean University, Shanghai 200090, China)
Abstract:【Objective】 Comparative study on morphological and skeletal differences of two Schizothorax species in Tarim River system. The provide scientific reference for the systematic evolution and geographical distribution pattern of the plateau fish.【Methods】 To study the external morphology and skeletal system of Schizothorax irregularis and Diptychus maculatus which are endemic to Xinjiang by anatomical comparison. Principal component analysis of morphological features was carried out by combining frame data and traditional quantifiable data and then the difference of bone morphology between CT scanning and boiling cutting was compared. 【Results】 Among 31 proportional traits of Schizothorax species, there were significant differences in 1-2/body length (BL) (Plt; 0.05), body width (BW/BL) and other 25 proportional traits were significantly different (Plt; 0.01); In the principal component analysis, the cumulative contribution rate of the first two principal components was 56.74%, and the difference was mainly reflected in the body head and the front torso. The number of cranial, pharyngeal and appendicular bones of the two Schizothorax species was the same, with 42, 80 and 132, respectively. However, there were some differences in the number of vertebrae and ribs between the two Schizothorax species: there were 4+42-43+1 vertebrae and 20-21 ribs in S. irregularis(2 tails), and there were 4+34-35+1 spines and 17-18 ribs in D. maculatus(2 tails). On some bones, such as the 2nd preethmoid, sphenotic, pterotic, premaxilla, maxilla, pterygoid and mstapterygoid, especially in the pharyngeal, the differences between the two Schizothorax species were more pronounced. 【Conclusion】 The morphological and skeletal differences of the two Schizothorax species in the Tarim River system are obvious, and their evolutionary mechanisms are closely related to the uplift of the Qinghai-Tibet Plateau. The unique morphological and evolutionary characteristics and skeletal differences can be further served as the basis for the classification and identification of fish, which might enrich the biological data of their morphological and evolutionary evolution.
Key words:Tarim River system; Schizothorax irregularis; Diptychus maculatus; morphological characteristics; skeletal anatomy
Fund projects:Special Financial Project of Ministry of Agriculture and Rural Affairs (Fishery Resources and Environment Survey in Key Waters of Northwest China); Science and Technology Public Relations Plan in Key Areas of XPCC Science and Technology Bureau \"Integration and Demonstration of Key Technologies for Comprehensive Development and Utilization of Saline-alkali Water Fishery in Southern Xinjiang\" (2022DB019); Key Laboratory Project of Tarim Animal Science and Technology of XPCC \"Research on Key Techniques of Artificial Breeding of Schizothorax pseudaksaiensis\" (HS201902)
Correspondence author: CHEN Shengao (1980-), male, from Xining, Qinghai, professor,Ph.D., doctoral supervisor, research direction: fishery resources and environment, (E-mail) chenshengao@163.com