
摘 要:【目的】研究新疆野生阿魏菇复合群的种群组成、地理分布和自然保有量等,为野生阿魏菇资源保育和开发利用提供理论支撑。
【方法】采用普查和样方调查相结合方法,采集新疆野生阿魏菇复合群资源,利用分子生物技术对收集菌株进行种类鉴定,运用统计学分析,阐明新疆野生阿魏菇复合群资源的种群组成、地理分布和自然保有量等。
【结果】新疆野生阿魏菇复合群资源由白灵侧耳和阿魏侧耳两个种群组成,白灵侧耳为优势种群,占鉴定总数97.23%,阿魏侧耳为弱势种群,占鉴定总数的2.77%;野生白灵侧耳种群广泛分布在新疆石河子市、托里县、裕民县、额敏县、福海县、富蕴县和青河县等7个区域,而野生阿魏侧耳种群仅分布在托里县、裕民县和额敏县等3个区域;野生白灵侧耳资源自然保有量总体呈下降趋势,且各分布区差异明显,其中石河子市、托里县和额敏县下降幅度最大,样方内年度采集数量最低年份只有0~1株,保育形势严峻;野生阿魏侧耳资源自然保有量极少,5年内在3个分布区样方内仅采集到3株野生菌株,保育形势尤为严峻。
【结论】新疆野生阿魏菇复合群由白灵侧耳和阿魏侧耳2个种群组成,白灵侧耳为优势种群,且为广布种;野生白灵侧耳和阿魏侧耳资源自然保有量总体均呈下降趋势。
关键词:阿魏菇复合群;白灵侧耳;阿魏侧耳;地理分布
中图分类号:S646;S188 文献标志码:A 文章编号:1001-4330(2024)04-0996-08
0 引 言
【研究意义】野生阿魏菇是一类寄生或腐生在伞形科植物阿魏(Ferula spp.)根茎上的侧耳属真菌的统称,其子实体具有很高的食药用价值[1, 2]。在我国野生阿魏菇资源仅分布于新疆戈壁或山地阿魏滩上,保有量稀少,因此野生阿魏菇分类学地位和物种组成是一个复杂的问题[5]。新疆阿魏菇自然群体是由形态难辨的白灵侧耳(Pleurotus tuoliensis)和阿魏侧耳(P. eryngii var. ferulae)组成的复合群[6]。收集野生阿魏菇样本并鉴定,评价研究野生阿魏菇复合群种群组成、地理分布及自然保有量等,对新疆野生阿魏菇种质资源开发利用和保育保护研究具有重要意义。【前人研究进展】野生阿魏菇的分布区域最早报道在新疆伊犁哈萨克自治州(简称伊犁州)、塔城地区、阿勒泰地区和昌吉回族自治州(简称昌吉州)木垒县等地[4, 7],其中尤以塔城地区报道最多,且塔城地区野生阿魏菇资源具有丰富的遗传多样性[8]。贾培松等[9]报道野生阿魏菇资源在石河子市也有分布,并对该区域阿魏菇资源遗传多样性进行了评价研究。牟川静[10]发现白灵侧耳种群后,先后被鉴定命名为阿魏侧耳托里变种(P. eryngii var. tuoliensis)、内布罗迪侧耳(P. nebrodensis)[11]、刺芹侧耳托里变种(P. eryngii var. tuoliensis)[12, 13]和刺芹侧耳托里亚种(P. eryngii subsp. tuoliensis)[14]。Zhao et al.基于形态学和多基因分子系统学研究认为刺芹侧耳托里亚种应为刺芹侧耳种族群的一个独立物种,将其分类地位从亚种提升为种级分类单元,命名为白灵侧耳(P. tuoliensis)[15]。Fu et al.(2017)和Zhang et al.(2018)[16, 17]基于分子标记和基因组学研究,也支持白灵侧耳为一个独立于P. eryngii的种的论断。2018年使用规范名称为白灵侧耳P. tuoliensis (C.J. Mou) M.R. Zhao amp;Jin X. Zhang,阿魏侧耳P. eryngii var. ferulae (Lanzi) Sacc. [18]。我国新疆地区阿魏菇自然种群为复合群,包括白灵侧耳(P. tuoliensis)和阿魏侧耳(P. eryngii var.ferulae)。【本研究切入点】虽然前人研究报道了新疆野生阿魏菇资源在地理分布、种群组成等阶段性研究结果,但由于样本量偏少、覆盖区域有限,不能代表野生阿魏菇资源的整体情况。虽然前期研究已经明确了野生阿魏菇为复合群及其部分分布区域,但前期研究使用的野生菌株数量和来源地均很有限,是否还有其他刺芹侧耳种族群、复合群各种群资源的具体地理分布、自然保有量等研究一直未见报道。
需研究野生阿魏菇所有分布区域和更大规模鉴定分析野生阿魏菇样本,阐明野生阿魏菇复合群种群组成、地理分布及自然保有量等。【拟解决的关键问题】采用普查和样方调查相结合方法,采集新疆野生阿魏菇复合群资源为研究对象,利用分子生物技术对收集菌株进行种类鉴定,通过统计学分析,阐明新疆野生阿魏菇复合群资源的种群组成、地理分布和自然保有量,为野生阿魏菇资源保育和开发利用提供理论支撑。
1 材料与方法
1.1 材 料
1.1.1 野生阿魏菇
以2011年至2022年间收集的野生阿魏菇复合群资源为材料,野生菌株经子实体组织分离,纯化培养后获得菌丝体纯培养物,母种保存于新疆农业科学院植物保护研究所种质资源库。
1.1.2 供试培养基
PDA(Potato Dextrose Agar)培养基:马铃薯200 g,葡萄糖20 g,琼脂粉20 g,补水至1 000 mL,pH自然。
1.2 方 法
1.2.1 野生阿魏菇复合群资源
1.2.1.1 调查区域
选择新疆石河子市、塔城地区、阿勒泰地区、昌吉州和伊犁州的10个县(市)作为调查区域。采用随机踏查法对10个区域阿魏滩上的野生阿魏菇资源进行大范围的调查和采集,统计发现的所有野生阿魏菇子实体。
1.2.1.2 样方调查
对普查中有野生阿魏菇资源分布的区域进行连续5年的样方调查,每个采集区随机划定5个样点,每个样点踏查面积为50 m×50 m,每个样点均采用地毯式踏查方式,所有野生阿魏菇子实体。同时收集野生阿魏菇分布区的地理信息,包括地形、植被类型、地理坐标和海拔等。
对于符合采集要求的野生阿魏菇子实体,利用组织分离法对采集的子实体进行菌种分离,获得野生阿魏菇菌株的菌种,经纯化培养后获得菌丝体纯培养物,于4℃保藏箱保藏。
1.2.2 野生阿魏菇菌株鉴定
1.2.2.1 菌丝体培养与收集
取出保藏的阿魏菇菌种,在无菌条件下取黄豆粒大小菌种块接种至PDA培养基平板上,于25℃培养箱黑暗培养7~10 d,使用打孔器沿菌落边缘取相同菌龄直径为5 mm的菌饼接种至铺有玻璃纸的PDA平板中央,于25℃培养箱黑暗培养。待菌丝体长满平板时收集菌丝体,用于DNA提取。
1.2.2.2 基因组DNA提取
利用试剂盒法提取阿魏菇菌株DNA,通过1%琼脂糖凝胶电泳检测DNA的质量和浓度,将合格的DNA保存于-20℃或-80℃冰箱保存,用于PCR。
1.2.2.3 分子鉴定
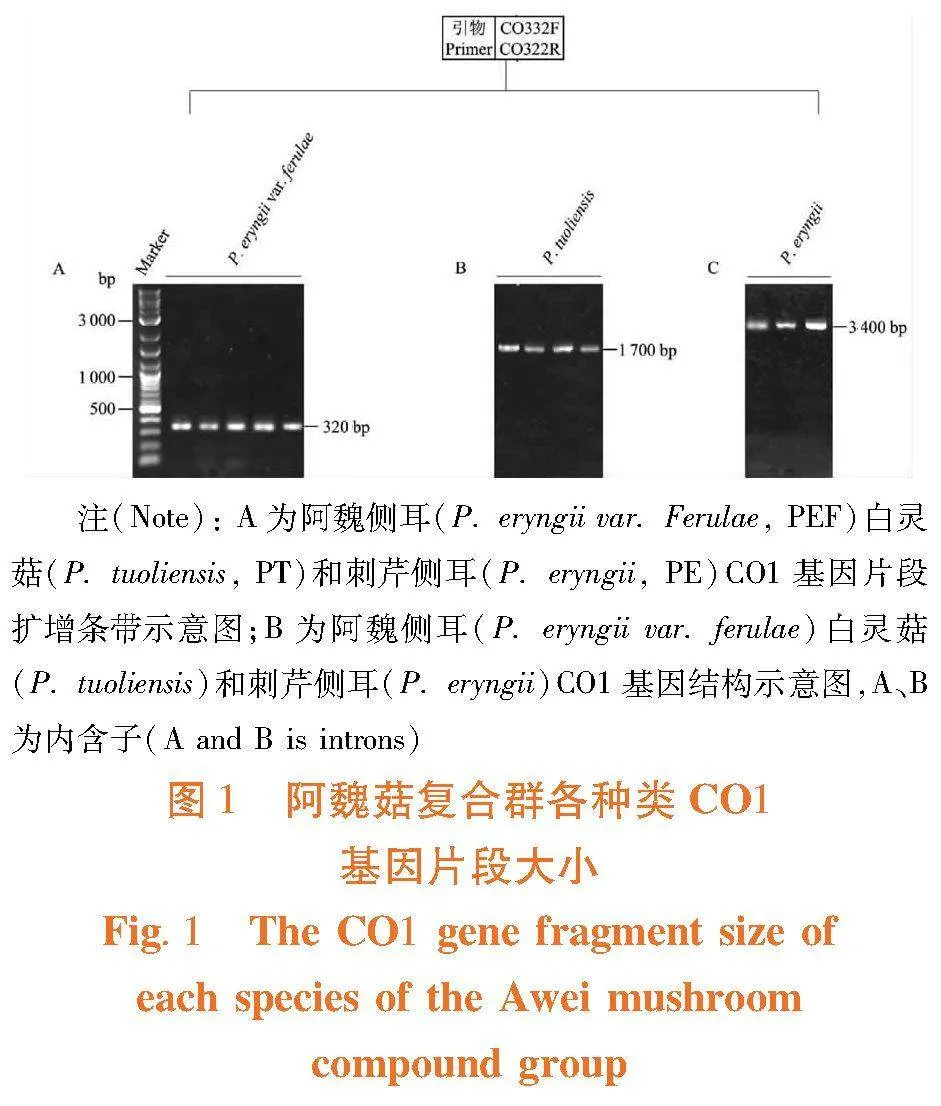
利用CO1基因片段对阿魏菇菌株进行种类鉴定,CO1基因扩增引物:CO332F:5′-TTACAAGGAGATCATCAATT-3′;CO332R:5′-TGCTAAGTGTAGACTGAAGA-3′。PCR 反应体系(25 μL):r Taq酶(5 U/μL)0.1 μL,dNTPs(各2.5 mmol)1 μL,10×r Taq buffer (Mg2+ plus)2.5 μL ,CO332F(10μmol)1 μL,CO332R(10μmol)1 μL,基因组DNA 2 μL,ddH2O 17.4 μL。PCR 扩增程序:94℃预变性5 min;然后94℃变性40 s,53℃退火40 s,72℃延伸120 s,32 个反应循环;最后72℃补平7 min,4℃保存。
扩增产物经1.0%的琼脂糖凝胶电泳检测,根据片段大小即可将白灵侧耳(P. tuoliensis)、阿魏侧耳(P. eryngii var. ferulae)和刺芹侧耳(P. eryngii)等分开,白灵侧耳PCR片段大小为1 700 bp,阿魏侧耳PCR片段大小为320bp,刺芹侧耳PCR片段大小为3 400 bp[19]。图1
1.3 数据处理
应用Excel 2010软件对数据进行数据处理、统计学分析。
2 结果与分析
2.1 野生阿魏菇复合群组成
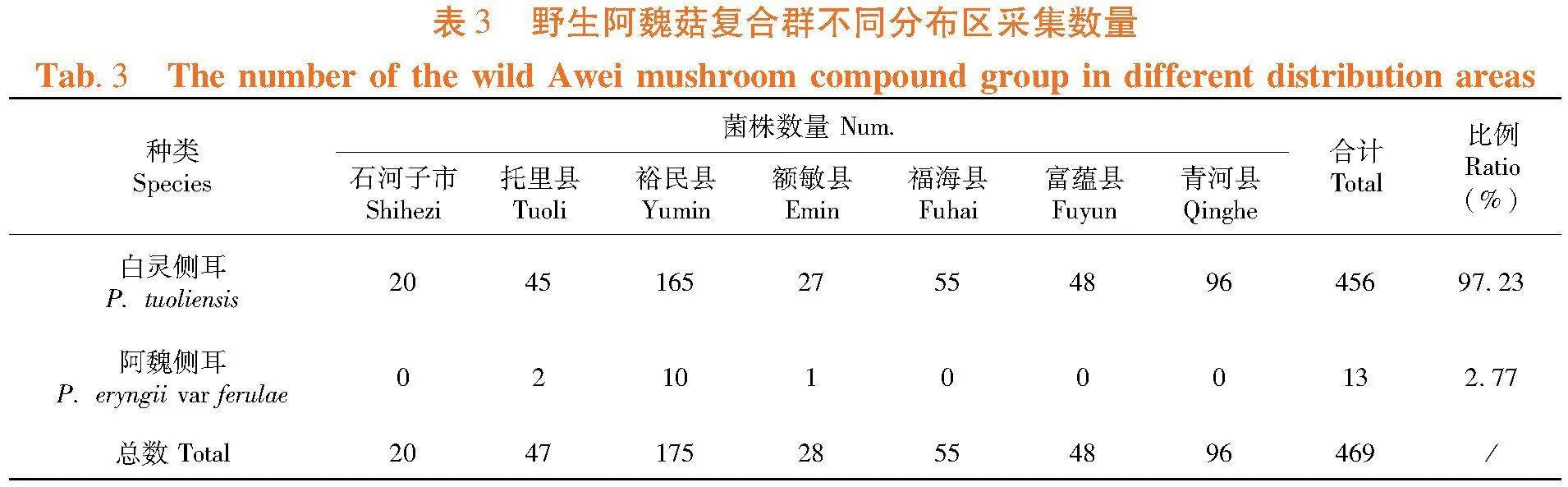
研究表明,469个野生菌株中,共鉴定到白灵侧耳菌株456株,鉴定到阿魏侧耳菌株13株,分别占鉴定总数的97.23%和2.77%;从年度种群组成看,野生白灵侧耳菌株数量占比在50%~100%,而野生阿魏侧耳在0%~50%,平均年度数量不足2株,新疆野生阿魏菇复合群由白灵侧耳和阿魏侧耳两个种群构成,白灵侧耳为优势种群,阿魏侧耳为弱势种群。表1
2.2 野生阿魏菇复合群分布区域
2.2.1 野生阿魏菇复合群资源整体集中分布区域
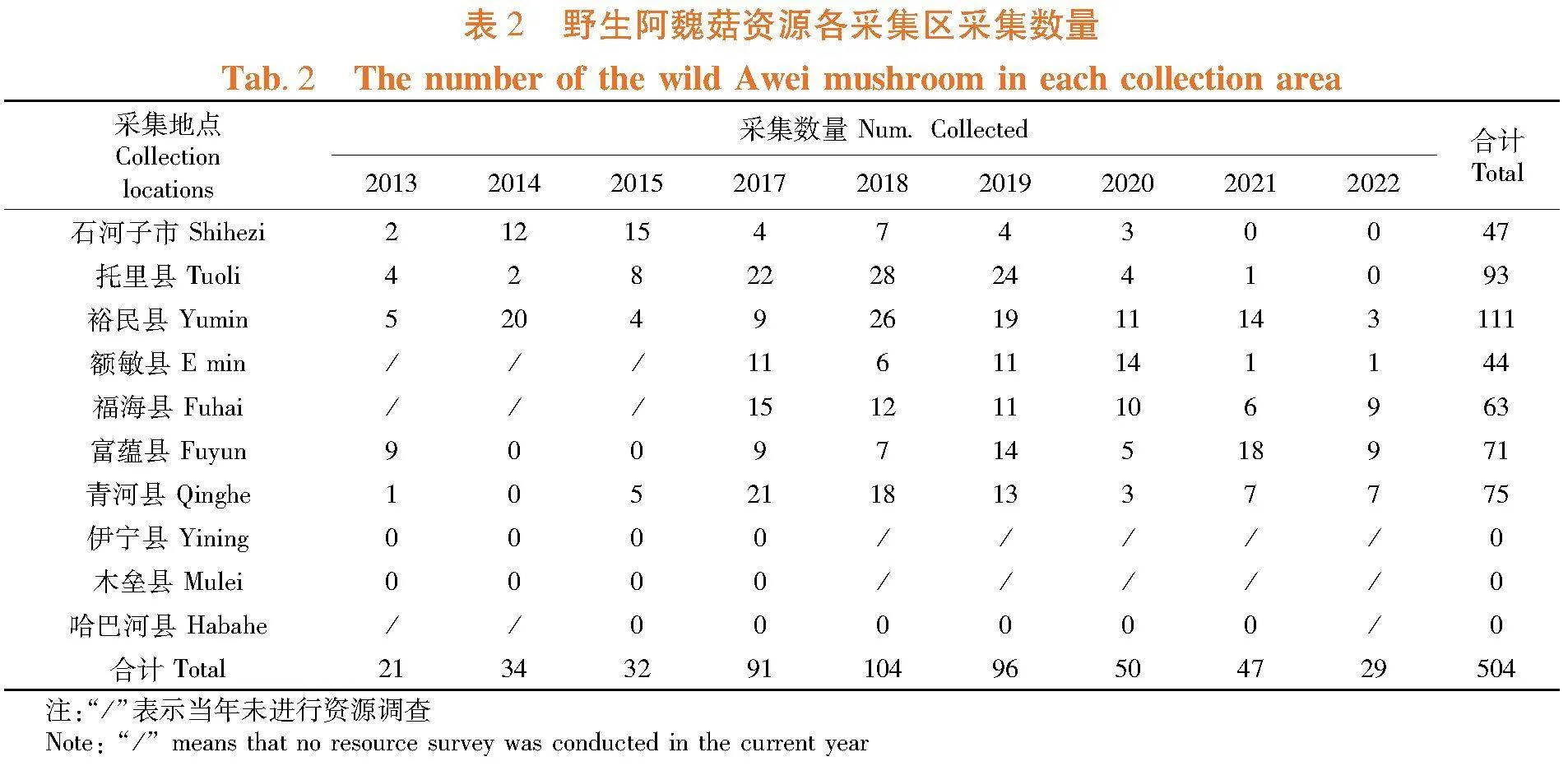
研究表明,10个野生阿魏菇资源采集区中,仅在石河子市、托里县、裕民县、额敏县、福海县、富蕴县和青河县等7个区域采集到野生阿魏菇资源,而在先前有野生阿魏菇资源报道的区域木垒县、伊宁县及哈巴河县未采集到野生阿魏菇资源。野生阿魏菇资源的生存区域有不断缩减的趋势,目前主要分布区域集中在新疆准噶尔盆地周边地区。表2
2.2.2 野生阿魏菇复合群不同种群分布区域
研究表明,野生白灵侧耳种群在石河子市、托里县、裕民县、额敏县、福海县、富蕴县和青河县等均有分布,在各区域收集到的野生菌株数量在20~165株;而野生阿魏侧耳种群仅在托里县、裕民县和额敏县等3个区域有分布,收集到的野生菌株数量稀少,分别为2株、10株和1株。野生白灵侧耳资源在新疆广泛分布,为优势种群,而野生阿魏侧耳资源分布区域有限,为弱势种群。表3
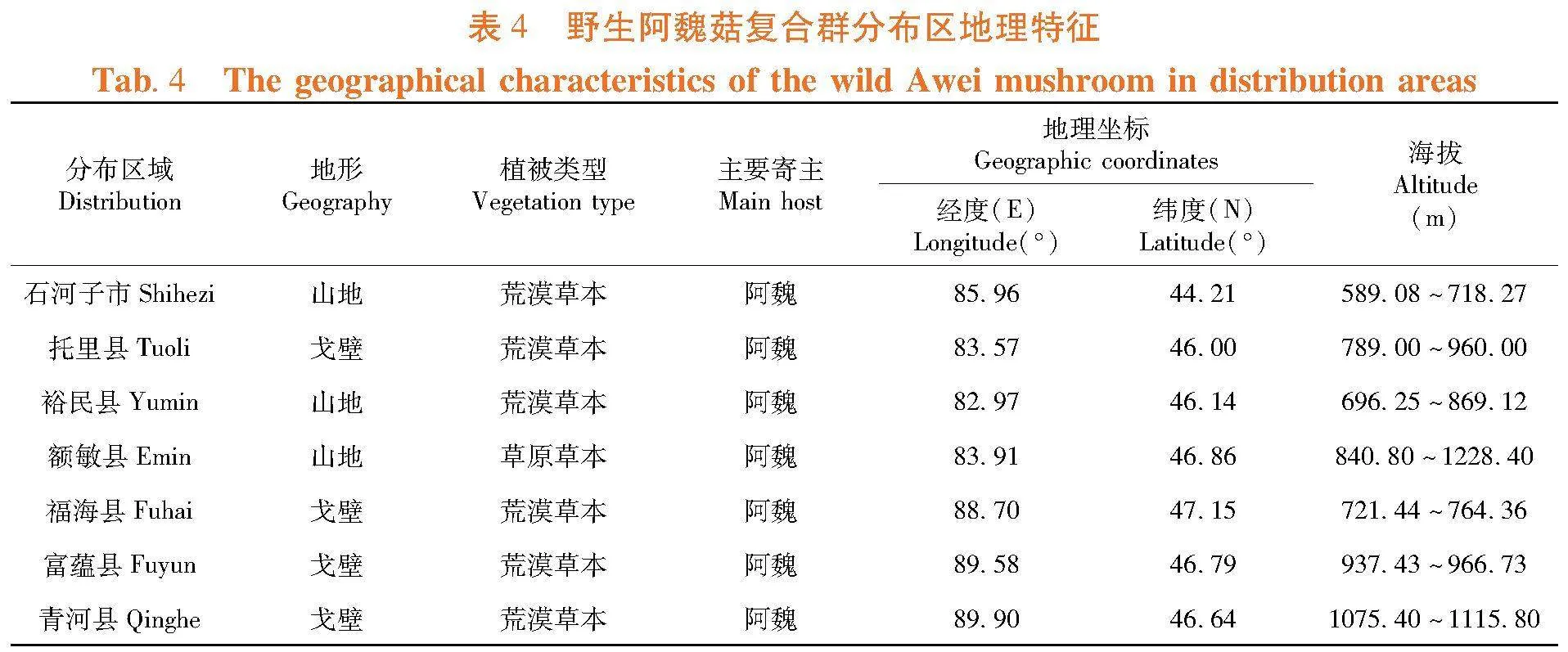
2.2.3 野生阿魏菇复合群分布区地理特征
研究表明,野生阿魏菇资源分布区地形地貌主要为山地和戈壁;各分布区的主要植被类型为荒漠草本植物,仅额敏县为草原草本植物;野生阿魏菇资源分布在82.97°~89.90°E,44.21°~47.15°N;野生阿魏菇资源在海拔589.08~1 228 m间均有分布,其中最低海拔在石河子市,最高海拔在额敏县。表4
2.3 野生阿魏菇复合群不同种群资源自然保有量
2.3.1 野生白灵侧耳资源自然保有量变化趋势
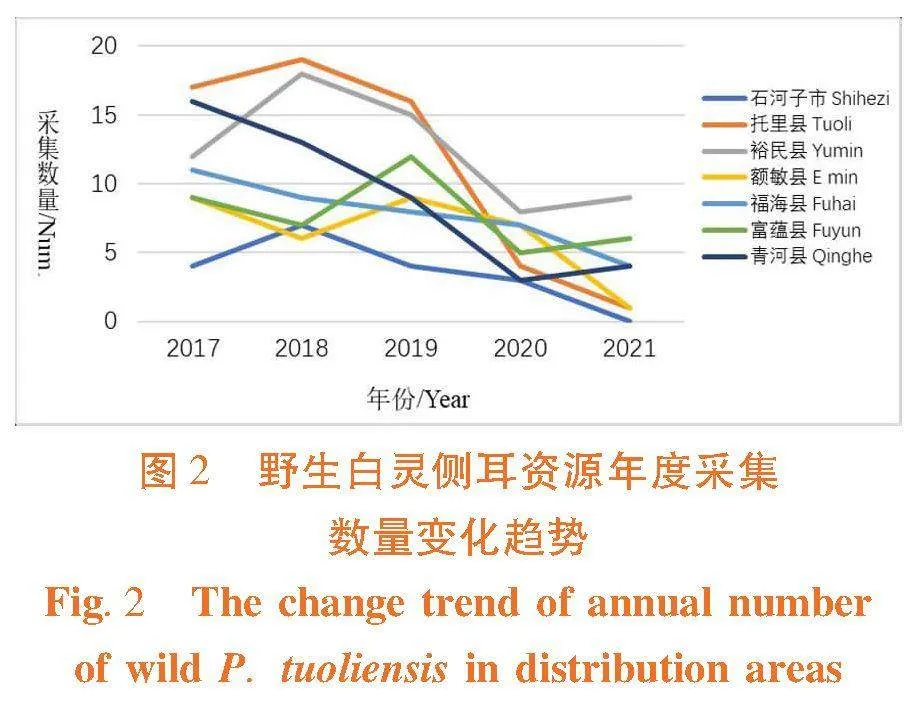
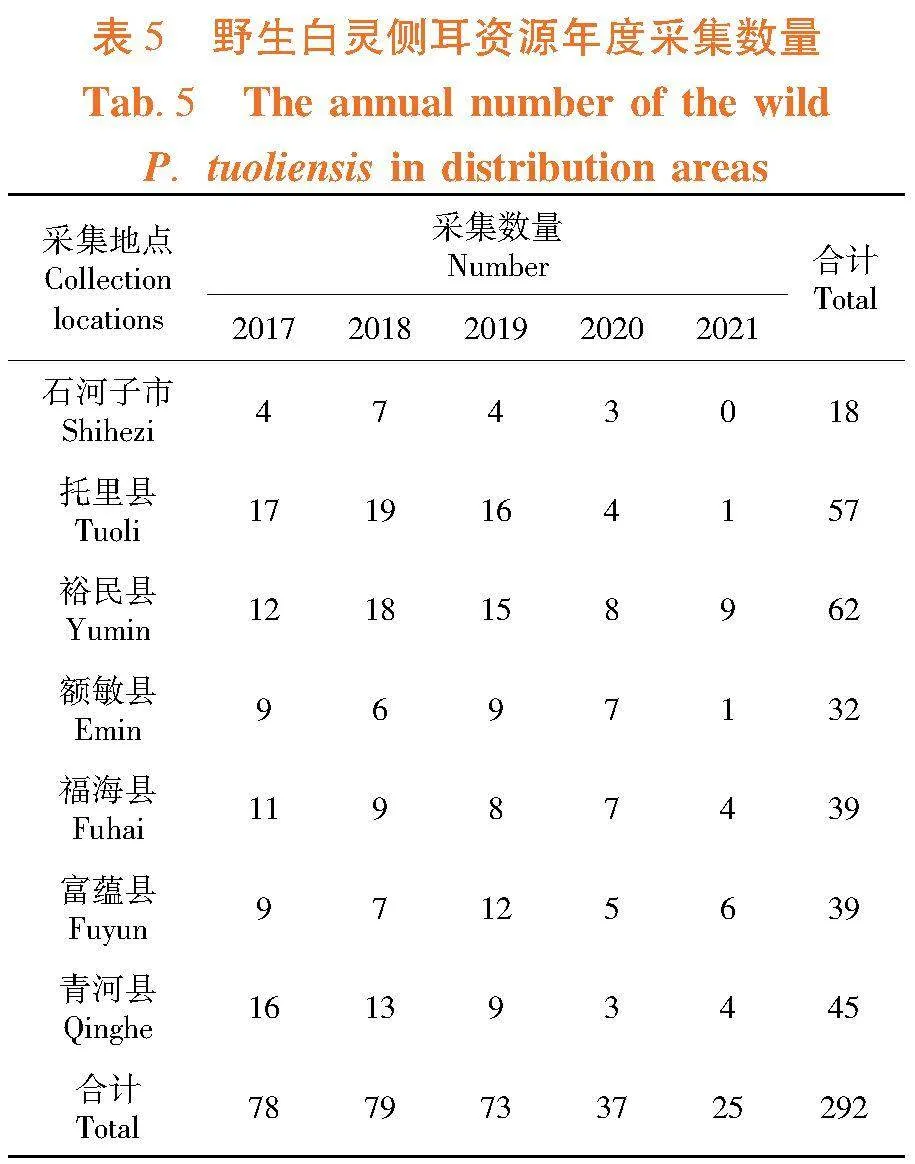
研究表明,野生白灵侧耳资源整体呈下降趋势,各分布区间下降趋势差异非常大,其中托里县、石河子市和额敏县下降幅度均较大,年度采集数量最多时分别为19、7和9株,而最低时(2021年度)采集数量分别为1、0和1株,下降幅度相对较小的区域为裕民县和富蕴县,年度采集数量最多时分别为18和12株,而最低时采集数量分别为8和5株;各地野生白灵侧耳资源数量差异较大,5年采集数量总和以裕民县最多,为62株,托里县次之,为57株,石河子最少,为18株。新疆区域野生白灵侧耳资源自然保有量呈逐年下降趋势,各分布区采集数量和下降幅度差异均非常大,其中石河子市、托里县和额敏县年度采集野生资源数量最少时仅为0~1株。表5,图2
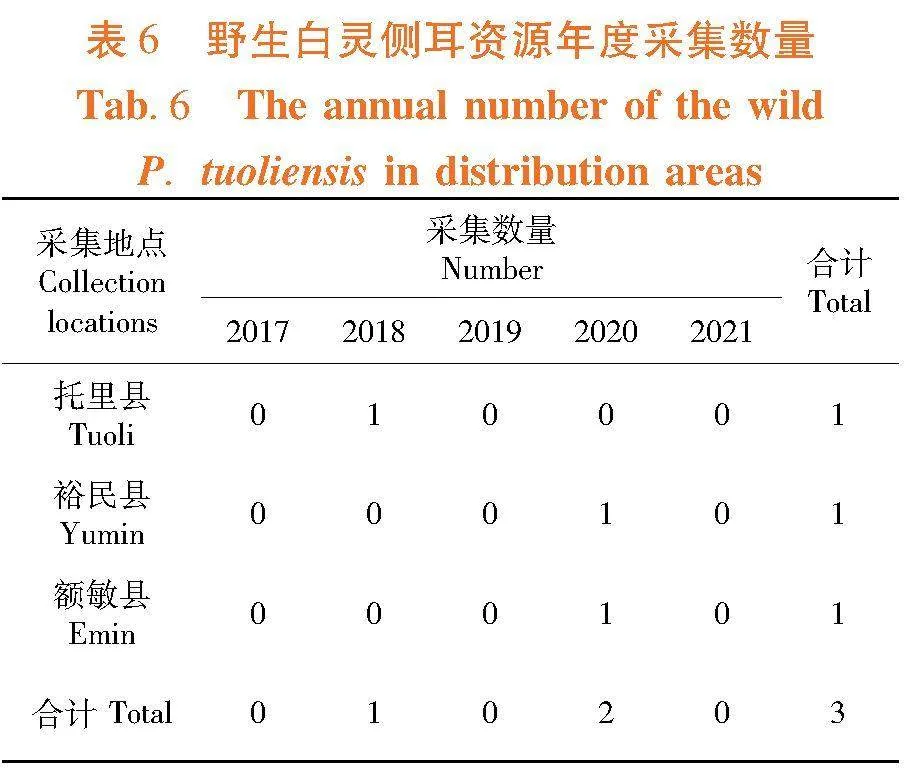
2.3.2 野生阿魏侧耳资源年度采集数量变化
研究表明,5年内3个野生阿魏侧耳资源分布区共采集到3株野生阿魏侧耳菌株,其中托里县于2018年度采集到1株,裕民县和额敏县分别于2020年度各采集到1株。新疆野生阿魏侧耳资源自然保有量十分稀少。表6
3 讨 论
野生阿魏菇自然群体是由形态难辨的白灵侧耳(Pleurotus tuoliensis)和阿魏侧耳(P. eryngii var. ferulae)组成的复合群[5]。研究对新疆范围内收集到的469株野生阿魏菇复合群菌株进行鉴定,也证实该复合群包括白灵侧耳和阿魏侧耳两个种群,暂未发现其他刺芹侧耳种群。
野生阿魏菇资源分布区多在戈壁阿魏滩和山地阿魏滩上,生存环境十分恶劣,自然保有量稀少,研究根据文献报道和调查情况选取了10个区域阿魏滩进行野生阿魏菇资源普查,结果显示野生阿魏菇资源分布在石河子市、托里县、裕民县、额敏县、福海县、富蕴县和青河县等7个区域,在先前报道有分布的木垒县和伊宁县阿魏滩,连续4年未发现野生阿魏菇,在哈巴河县也未发现该野生资源,野生阿魏菇资源自然保有量在不断降低,分布区在不断缩减。
4 结 论
新疆野生阿魏菇复合群由白灵侧耳和阿魏侧耳两个种群组成,白灵侧耳为优势种群,阿魏侧耳为弱势种群;野生白灵侧耳种群广泛分布在石河子市、托里县、裕民县、额敏县、福海县、富蕴县和青河县等7个区域,而野生阿魏侧耳种群仅分布在托里县、裕民县和额敏县等3个区域;野生白灵侧耳资源自然保有量较多,但总体呈下降趋势,且各分布区差异明显,部分区域在方内的年度采集数量最低年份只有1株或0株,保育形势严峻,而野生阿魏侧耳资源自然保有量极少,5年内在3个分布区采样点仅采集到3株野生菌株。
参考文献(References)
[1]陈忠纯. 我国阿魏侧耳的驯化与栽培[J]. 食用菌学报, 1996, 3(4): 11-14.CHEN Zhongchun. The domestication and cultivation of Pleurotus ferulae in China[J]. Acta Edulis Fungi, 1996, 3(4): 11-14.
[2] 贾培松, 罗影, 贾文捷, 等. 新疆野生阿魏蘑菌株主要农艺性状评价[J]. 新疆农业科学, 2018, 55(1): 134-142.JIA Peisong, LUO Ying, JIA Wenjie, et al. The evaluation of main agronomic traits of wild Pleurotus eryngii strains in Xinjiang[J]. Xinjiang Agricultural Sciences, 2018, 55(1): 134-142.
[3] 陈忠纯. 阿魏侧耳的研究[J]. 干旱区研究, 1991, 8(2): 94-95, 76.CHEN Zhongchun. Pleurotus ferulae lanzis research[J]. Arid Zone Research, 1991, 8(2): 94-95, 76.
[4] 曹玉清, 牟川静, 陈忠纯, 等. 阿魏侧耳某些生物学特性的初步研究[J]. 微生物学通报, 1985, 12(3): 97-101.CAO Yuqing, MU Chuanjing, CHEN Zhongchun, et al. Preliminary study on some biological characteristics of pleurotus ferulae lanzi[J]. Microbiology, 1985, 12(3): 97-101.
[5] Dai Y T, Sun L, Yin X L, et al. Pleurotus eryngii genomes reveal evolution and adaptation to the Gobi Desert environment[J]. Frontiers in Microbiology, 2019, 10: 2024.
[6] Zhang J X, Huang C Y, Ng T B, et al. Genetic polymorphism of Ferula mushroom growing on Ferula sinkiangensis[J]. Applied Microbiology and Biotechnology, 2006, 71(3): 304-309.
[7] 陈忠纯. 阿魏侧耳驯化初报[J]. 中国食用菌, 1986, 5(2): 16-17.CHEN Zhongchun. Preliminary report on domestication in pleurotus ferulae lanzi[J]. Edible Fungi of China, 1986, 5(2): 16-17.
[8] 贾培松, 努尔孜亚·亚力买买提, 罗影, 等. 新疆塔城地区野生阿魏菇菌株的遗传多样性分析[J]. 新疆农业科学, 2016, 53(10): 1900-1906.JIA Peisong,Nuerziya Yalimaimaiti,LUO Ying, et al. Key Laboratory of Integrated Management of Harmful Crop Vermin in China Northwestern Oasis, Minis, LUO Ying, et al. Genetic diversity analysis of wild P. eryngii strains from Tacheng, Xinjiang[J]. Xinjiang Agricultural Sciences, 2016, 53(10): 1900-1906.
[9] 贾培松, 努尔孜亚·亚力买买提, 管建华, 等. 新疆石河子地区野生阿魏菇菌株遗传多样性[J]. 西北农业学报, 2017, 26(1): 152-158.
JIA Peisong, Nuerziya Yalimaimaiti, GUAN Jianhua, et al. Genetic diversity of wild Ferula mushroom strains from Shihezi Area in Xinjiang[J]. Acta Agriculturae Boreali-occidentalis Sinica, 2017, 26(1): 152-158.
[10] 牟川静, 曹玉清, 马金莲. 阿魏侧耳一新变种及其培养特征[J]. 真菌学报, 1987, 6(3): 153-156.MOU Chuanjing, CAO Yuqing, MA Jinlian. A new variety of pleurotus eryngii and its cultural characters[J]. Mycosystema, 1987, 6(3): 153-156.
[11] 卯晓岚. 中国大型真菌[M]. 郑州: 河南科学技术出版社, 2000.MAO Xiaolan. The macrofungi in China[M]. Zhengzhou: Henan Science and Technology Press, 2000.
[12] Kawai G, Babasaki K, Neda H. Taxonomic position of a Chinese Pleurotus “Bai-Ling-Gu”: it belongs to Pleurotus eryngii (DC.: Fr.) Quél. and evolved independently in China[J]. Mycoscience, 2008, 49(1): 75-87.
[13] 黄晨阳, 陈强, 邓旺秋, 等. 中国栽培白灵菇学名的订正[J]. 植物遗传资源学报, 2011, 12(5): 825-827, 832.HUANG Chenyang, CHEN Qiang, DENG Wangqiu, et al. Correction of scientific Name for cultivated bai-ling-gu in China[J]. Journal of Plant Genetic Resources, 2011, 12(5): 825-827, 832.
[14] Zervakis G I, Ntougias S, Gargano M L, et al. A reappraisal of the Pleurotus eryngii complex–New species and taxonomic combinations based on the application of a polyphasic approach, and an identification key to Pleurotus taxa associated with Apiaceae plants[J]. Fungal Biology, 2014, 118(9/10): 814-834.
[15] Zhao M R, Huang C Y, Wu X L, et al. Genetic variation and population structure of the mushroom Pleurotus ferulae in China inferred from nuclear DNA analysis[J]. Journal of Integrative Agriculture, 2016, 15(10): 2237-2246.
[16] Fu Y P, Dai Y T, Yang C T, et al. Comparative transcriptome analysis identified candidate genes related to bailinggu mushroom formation and genetic markers for genetic analyses and breeding[J]. Scientific Reports, 2017, 7(1): 9266.
[17] Zhang Z B, Wen J W, Li J Z, et al. The evolution of genomic and epigenomic features in two Pleurotus fungi[J]. Scientific Reports, 2018, 8(1): 8313.
[18] 戴玉成, 杨祝良. 中国五种重要食用菌学名新注[J]. 菌物学报, 2018, 37(12): 1572-1577.DAI Yucheng, YANG Zhuliang. Notes on the nomenclature of five important edible fungi in China[J]. Mycosystema, 2018, 37(12): 1572-1577.
[19] 宋驰, 陈强, 徐璟煜, 等. CO1在侧耳属物种快速鉴定中的应用[J]. 菌物学报, 2011, 30(4): 663-668.SONG Chi, CHEN Qiang, XU Jingyu, et al. Application of CO1 for rapid identification of Pleurotus species[J]. Mycosystema, 2011, 30(4): 663-668.