![]()
doi:10.6048/j.issn.1001-4330.2024.05.020
摘" 要:【目的】比较两种酵母培养物及其代谢成分的差异,为指导酵母培养物的生产及应用提供参考。
【方法】材料为1种自研酵母培养物与达农威益康 XP酵母培养物,利用非靶标代谢组学UHPLC-QTOF-MS技术,分析比较二者代谢产物成分及差异。
【结果】(1)在二级类别注释下,正、负离子模式分别注释到614、497个化合物,主要代谢类别为有机酸,核苷、核苷酸及其衍生物,氨基酸及其衍生物,两种酵母培养物均无特有代谢成分,只在含量上有显著(Plt;0.05)差异。(2)共237个差异代谢物在正离子模式检出,176个显著(Plt;0.05)上调表达,61个显著(Plt;0.05)下调表达;136个差异代谢物在负离子模式检出,64个显著(Plt;0.05)上调表达,72个显著(Plt;0.05)下调表达。(3)差异代谢物的 KEGG(Kyoto Encyclopedia of Genes and Genomes)富集分析主要集中在丙氨酸、天冬氨酸和谷氨酸等代谢通路。
【结论】自研酵母培养物与达农威益康XP的代谢产物含量存在显著差异,但其发酵原料及工艺更简便,并含有多种具药理、生理作用的代谢成分,有潜在应用价值。
关键词:酵母培养物;非靶标代谢组学;差异代谢物;酿酒酵母
中图分类号:S188""" 文献标志码:A""" 文章编号:1001-4330(2024)05-1218-09
收稿日期(Received):
2023-10-15
基金项目:
新疆维吾尔自治区自然科学基金重点项目(2023D01D11);新疆生产建设兵团重点领域科技攻关计划(2022AB013);新疆生产建设兵团重点领域科技攻关计划(2020AB013)
作者简介:
林青(1985-),女,山东烟台人,副研究员,研究方向为微生物生态,(E-mail)qinglinxj@163.com
通讯作者:
娄恺(1968-),男,河南人,研究员,博士,硕士生导师,研究方向为微生物生态,(E-mail)loukai@tsinghua.org.cn
霍向东(1974-),男,甘肃人,副研究员,博士,研究方向为微生物资源,(E-mail)xiangdonghuo@163.com
0" 引 言
【研究意义】酵母培养物(Yeast Culture,YC )是一种由酵母及其生长培养基组成的酵母发酵产物[1],具有无毒无污染、营养物质丰富、提高动物生产性能和免疫力等特点[2]。酵母培养物是一类非标产品,我国目前尚未出台相关国家标准,因此市场现有产品质量良莠不齐。【前人研究进展】潘韵等[3]研究了7种酵母培养物的成分和对仔猪的应用效果,结果表明3个产品的质量标准、风味和颜色及应用效果显著优于其他产品,益康XP酵母培养物在奶牛泌乳[4]、母猪繁殖[5]、鸡产蛋[6]、断奶仔猪[7]和羔羊生长[8]性能等方面均有显著效果。有研究将益康XP作为对照,进行酵母培养物的有效组分分析[9]。【本研究切入点】酵母培养物成分复杂,除能检测到的成分外,仍有许多“未知营养生长因子”在发挥作用[10]。而且,酵母培养物的成分和性能还因不同生产菌种、培养基成分和发酵工艺条件等因素产生巨大差异[11],需进一步探究其作用机理的复杂性。【拟解决的关键问题】代谢组学是研究有机体及细胞内各种低分子量代谢物组成和动态变化,反映机体生理代谢状态,揭示在遗传因素和环境变化影响下,代谢物发生的变化规律和相关反应机制[12-14]。利用非靶标代谢组学技术比较差异代谢物,分析二者成分特点及优势代谢产物,为了解酵母培养物发酵特性及酵母培养物的生产与应用提供参考。
1" 材料与方法
1.1" 材 料
菌种:菌株Saccharomyces cerevisiae来自新疆农业科学院微生物应用研究所。
原料:玉米粉、麸皮、豆粕、糖蜜、糖化酶(100 U/g)、淀粉酶(200 U/g)由新疆天康饲料科技有限公司生物添加剂分公司提供。达农威益康 XP 酵母培养物购自达农威生物发酵工程技术(深圳)有限公司。
试剂:甲醇、乙酸铵、乙腈、氨水(色谱纯),吡啶、氯仿(色谱纯),甲氧铵盐(分析纯),BSTFA(含 1% TMCS, v/v),L-2-氯苯丙氨酸(纯度≥98%),标准品:L-2-氯苯丙氨酸,饱和脂肪酸甲酯(纯度≥98%)。
仪器:气相色谱(7890-B),色谱柱(DB-5MS),超高效液相色谱(1290-UHPLC),质谱仪(PEGASUS HT),高分辨质谱(Triple TOF-6600),超声仪(YM-080S),离心机(Heraeus Fresco-17),烘箱(DHG-9023A),超低温冰箱(Forma-900 series),分析天平(BSA-124S-CW),真空干燥仪(LNG-T98)研磨仪(JXFSTPRP-24)。
1.2" 方 法
1.2.1" 酵母培养物的制备
菌种活化:将冻干保藏的菌株Saccharomyces cerevisiae在PDA固体培养基活化,28℃培养48 h。
液体发酵:将活化好菌株接种于麦芽汁液体培养基,200 r/min,28℃培养24 h,作为种子液按5%接种量接种于麦芽汁液体培养基,200 r/min,28℃培养24 h。
固态发酵:调配玉米粉、麸皮、豆粕、糖蜜成碳氮比为1∶16固体发酵底物,将上述液体发酵物按20%接种量接入固体发酵底物,同时添加糖化酶100 U/g和淀粉酶200 U/g,充分混匀,30℃厌氧发酵3 d,50℃烘干后,液氮速冻 15 min,-80℃保存。将自研酵母培养物设置为A组,达农威益康 XP 酵母培养物设置为B组,每组设置3个平行处理,采用 UHPLC-QTOF-MS技术,比较两种酵母培养物的成分组成和代谢物差异。代谢物的提取及后续分析均委托上海阿趣生物科技有限公司完成。
参考鲁晓宇等[15]方法配制,代谢物提取和色谱质谱参数设定。
1.3" 数据处理
使用ProteoWizard 3.0软件将原始数据格式转换为mzXML。峰数据处理后,鉴定峰的物质。使用 KEGG数据库注释鉴定到的代谢物。使用SIMCA 13.0软件,对数据进行对数、中心化处理,进行主成分分析(principal component analysis,PCA)和偏最小二乘法判别分析(partial least square discriminant analysis,PLS-DA),获得代谢物的VIP 值。使用 SPSS 20.0 软件统计分析, 计算各代谢物显著性(P 值)和差异倍数(FC 值)。差异代谢物的筛选设定阈值为 VIPgt;1.0,P lt;0.01,FC(A/B)gt; 2 或lt; 0.5,MS2 scoregt;0.9。
2" 结果与分析
2.1" 酵母培养物中代谢物成分差异比较
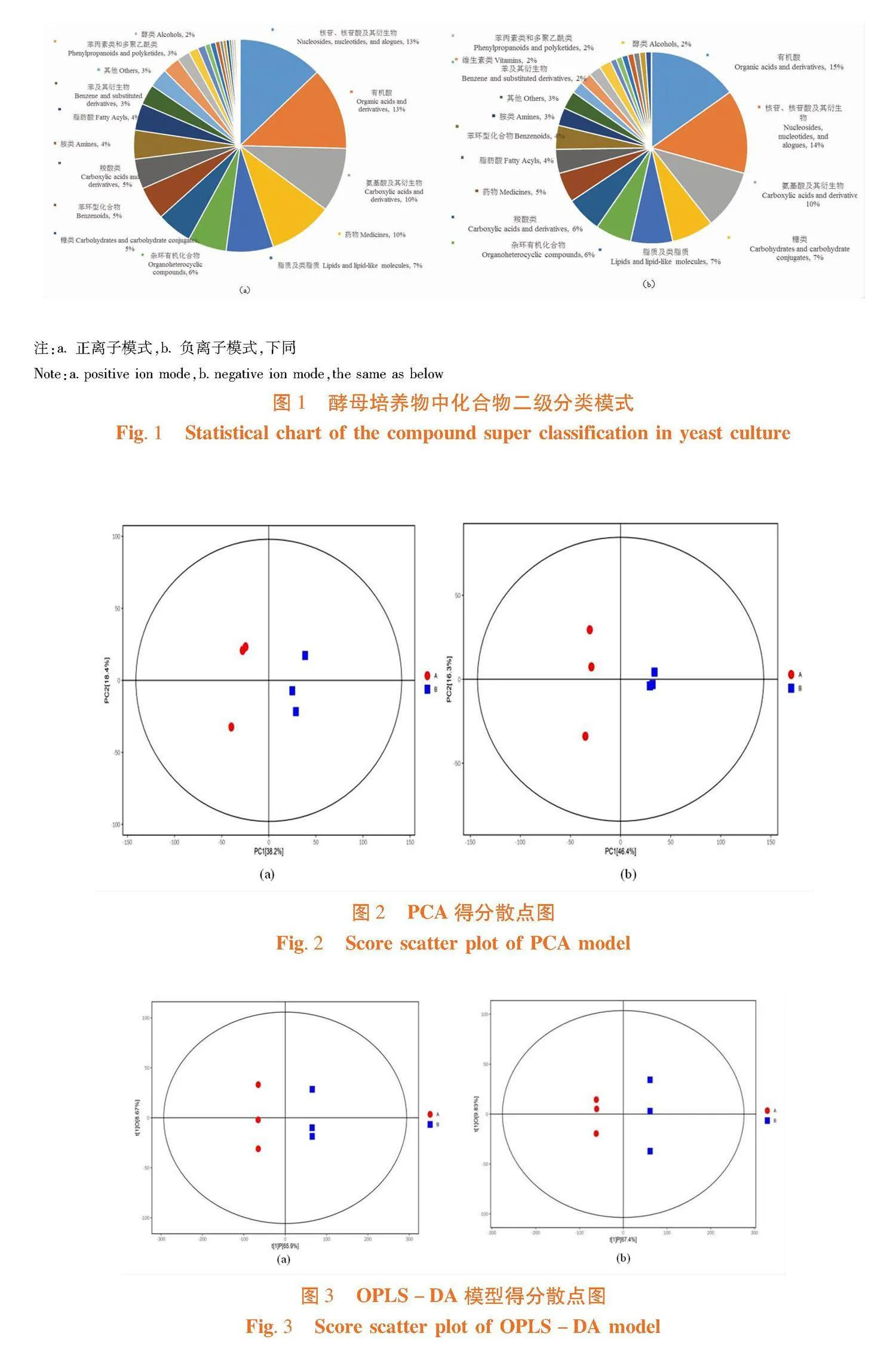
研究表明,在正离子和负离子模式下分别筛选出7 752和 6 709 个化合物,通过与数据库匹配,分别注释到4 940 和4 511 个化合物。正离子模式下,前614个化合物分类为30个二级类别,核苷、核苷酸及其衍生物(13%),有机酸(13%),氨基酸及其衍生物(10%),药物(10%)等为占比10%以上的主要类别。负离子模式下,前497个化合物分类为30个二级类别,有机酸(15%),核苷、核苷酸及其衍生物(14%),氨基酸及其衍生物(10%)等为占比10%以上的主要类别。两种酵母培养物代谢产物丰富,主要代谢类别为有机酸,核苷、核苷酸及其衍生物,氨基酸及其衍生物,在二级类别注释下,两种酵母培养物均无特有代谢物成分,代谢产物只在含量比例上有所差异。图1
2.2" 两种酵母培养物的PCA和PLS-DA分析
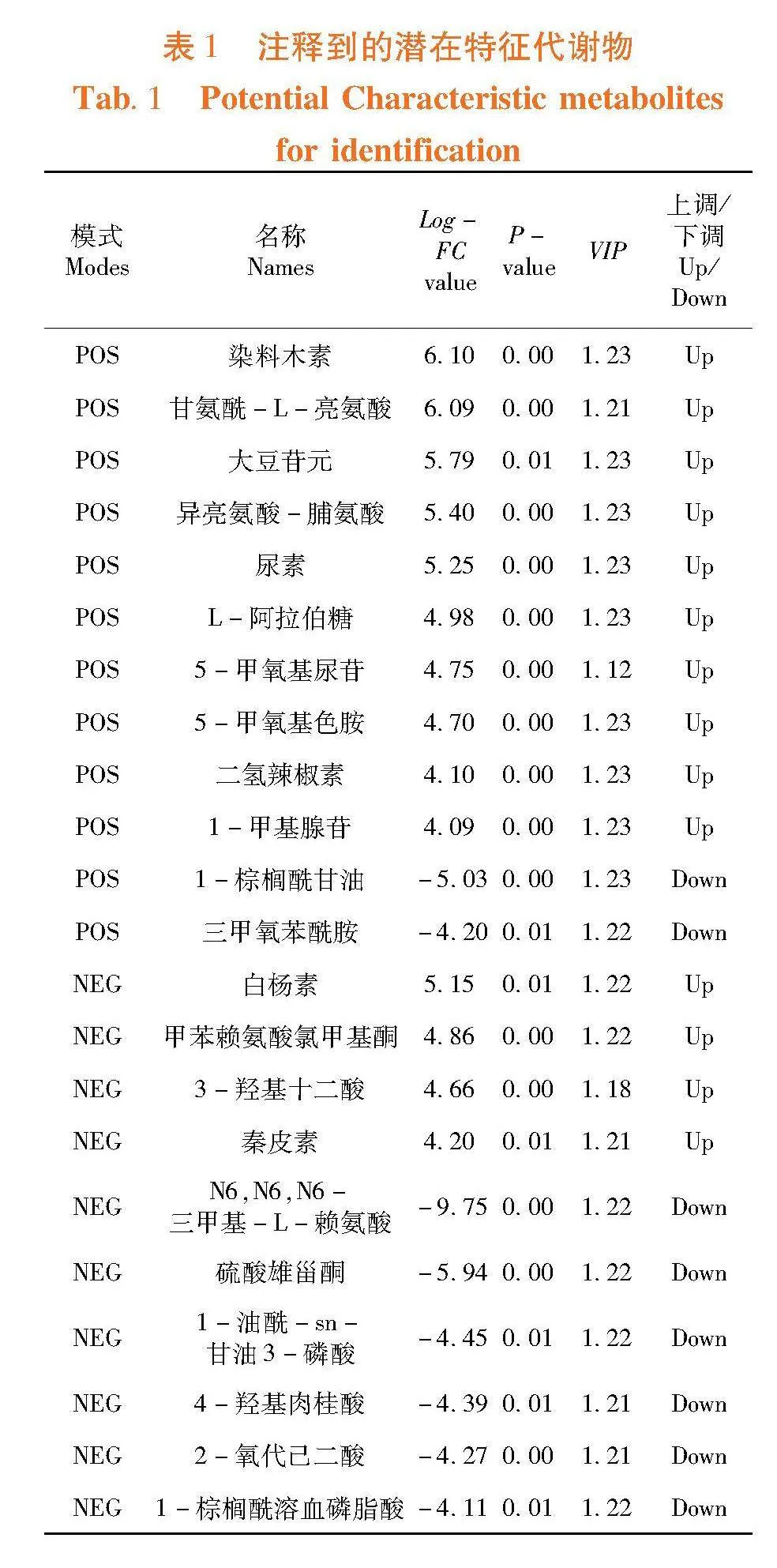
研究表明,从两种酵母培养物的6个样本中,共保留了14 461个有效峰。在正离子和负离子模式下,A组与B组组间分离明显,组内平行性好,2组样本具有不同的代谢谱。PCA模型中,样本均在95%置信区间,正离子、负离子模式下,前2个主成分的R2 X分别为0.566和0.627,模型对X变量解释能力较好。图2
R2 X、R2 Y是模型分别对 X、Y 矩阵的解释率,Q2表示模型预测能力。样本全部处于95%置信区间内,两种酵母培养物聚类分离效果非常显著,划分在左右两个象限内。正离子模型中,R2 X=0.746,R2 Y=1.000,Q2 =0.990,负离子模型中,R2 X=0.772,R2 Y=1.000,Q2 =0.992,两个模型中3个参数数值均接近于1 ,模型的预测能力及拟合准确性较高。图3
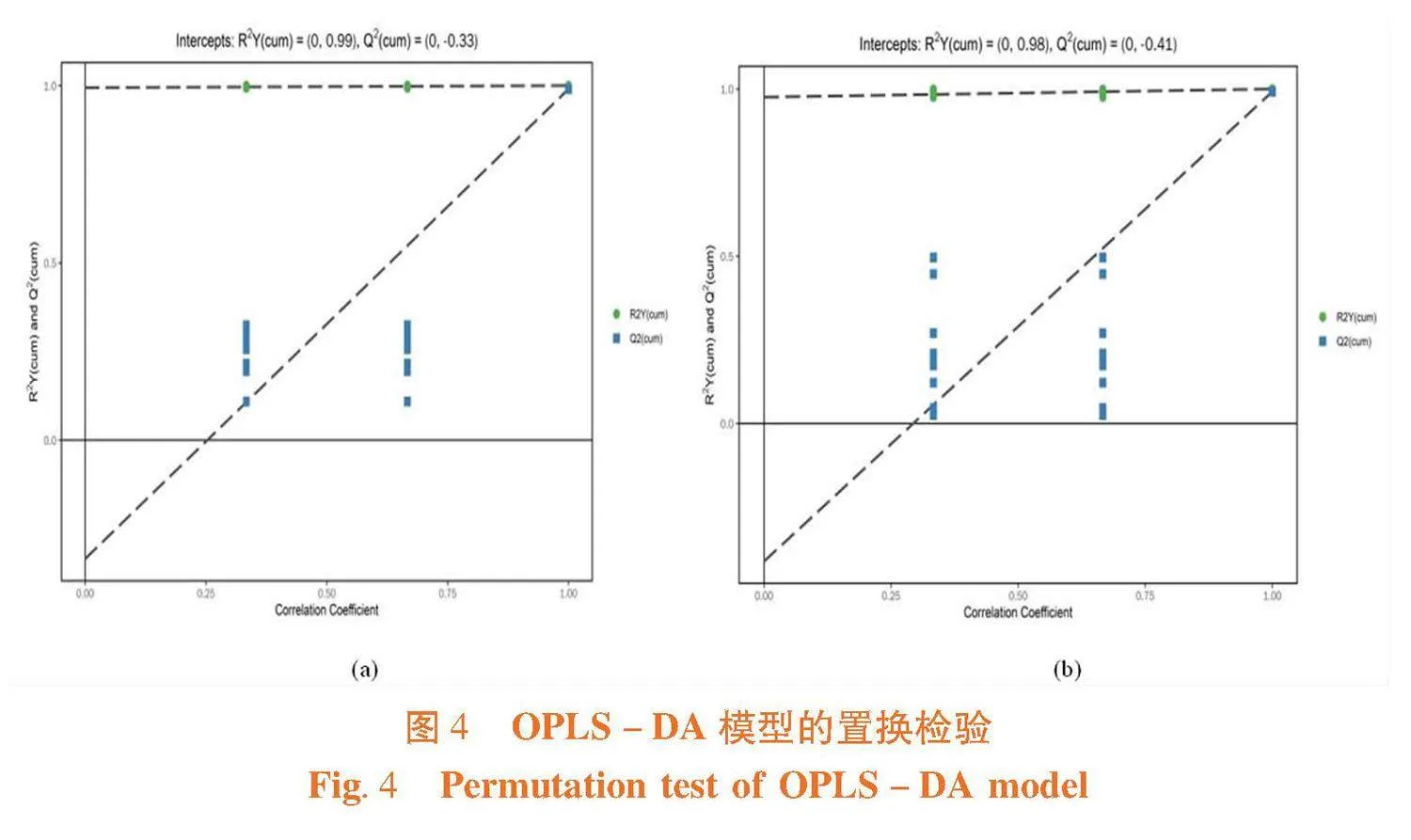
两种离子模式下,原模型的R2Y值和Q2值接近1,建立模型可以反映样本数据的真实情况,原模型解释A、B组间差异较好。置换检验结果随机模型Q2lt;原模型Q2,两种离子模式下,Q2回归线的截距分别为-0.33和-0.41,原模型稳健性良好,无过拟合现象,具有统计学意义,两种酵母培养物的代谢物存在显著差异。图4
2.3" 差异代谢物的筛选与鉴定
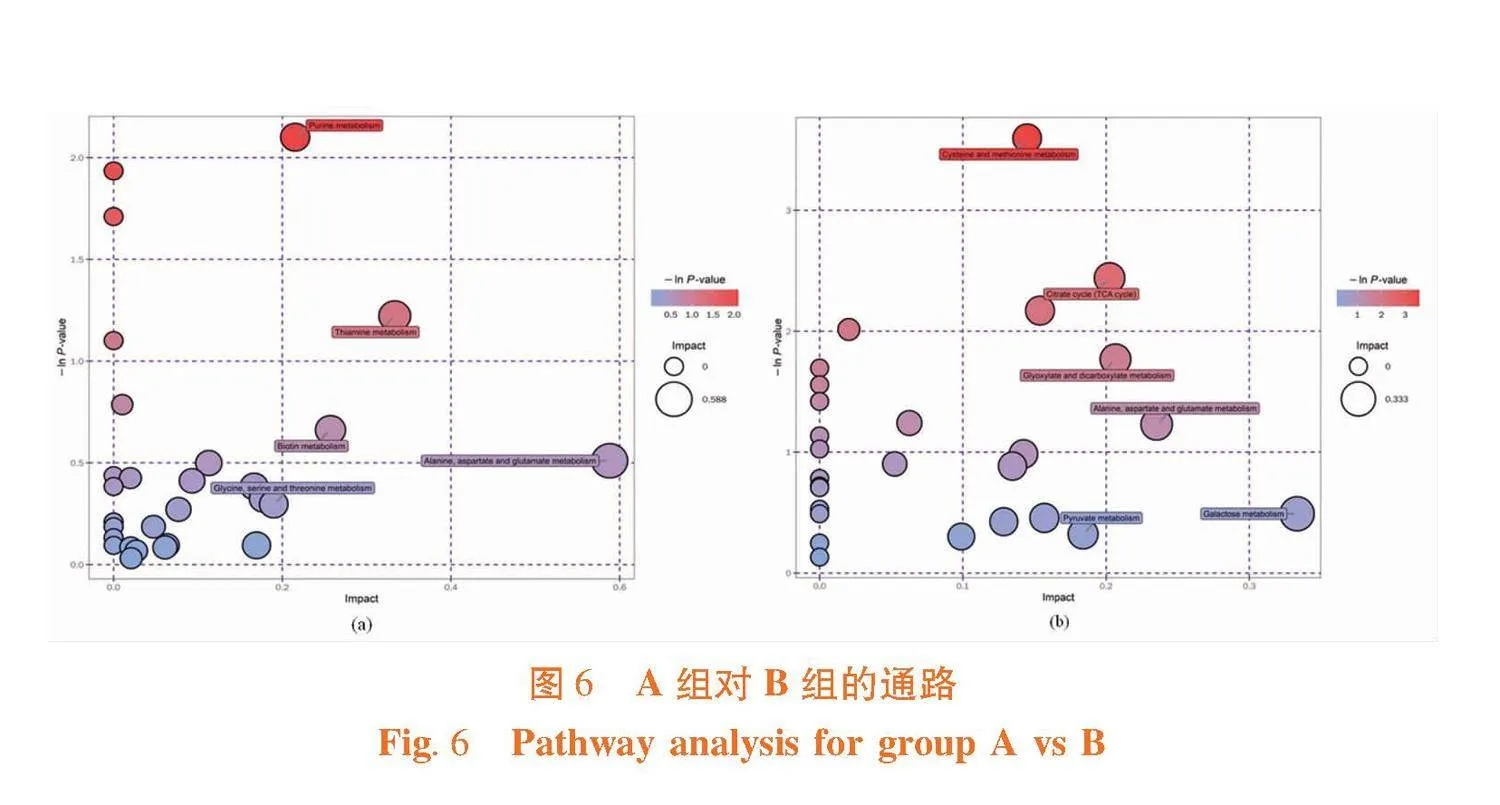
研究表明,正离子模式中,在两种酵母培养物中共检测到 237 种差异代谢物,有 176 种显著上调表达,包括染料木素、甘氨酰-L-亮氨酸、大豆苷元等,有 61 种显著下调表达,包括1-棕榈酰甘油、三甲氧苯酰胺等;负离子模式中,检测到 136 种差异代谢物,显著上调表达的有 64 种,包括白杨素、甲苯赖氨酸氯甲基酮等,显著下调表达的有72种,包括N6,N6,N6-三甲基-L-赖氨酸、硫酸雄甾酮等。两种离子模式下,A组样本共有240种代谢物显著上调,B组样本共有133种代谢物显著上调。图5
正负离子模式下,log FC值gt;4的代谢物共有14种,自研酵母培养物比达农威酵母培养物中代谢产物相对含量高于16倍以上的成分,其中染料木素log FC最大值为6.10,在自研酵母培养物中的含量是达农威酵母培养物的68.59倍。log FC值lt;-4的代谢物共有8种,达农威酵母培养物比实验组酵母培养物中代谢产物相对含量高于16倍以上的成分,其中N6,N6,N6-三甲基-L-赖氨酸log FC最大值为-9.75,在达农威酵母培养物中的含量是自研酵母培养物的861.08倍。表1
2.4" 差异代谢物 KEGG 富集通路
研究表明,正离子模式中,两组酵母培养物差异代谢物主要富集在丙氨酸、天冬氨酸和谷氨酸代谢通路,硫胺素代谢通路,生物素代谢通路,嘌呤代谢通路,甘氨酸、丝氨酸和苏氨酸代谢通路等;负离子模式中,主要注释和富集在半乳糖代谢通路,丙氨酸、天冬氨酸和谷氨酸代谢通路,乙醛酸和二羧酸代谢通路,柠檬酸循环,丙酮酸代谢通路,半胱氨酸和蛋氨酸代谢通路等。图6
3" 讨 论
3.1
采用多元变量统计分析方法,减少数据维度[16]。酵母培养物是成分复杂的发酵类产品[17],采用高效液相色谱检测益康XP酵母培养物含有上百种代谢产物,除检测出的营养物质外,还有许多未知成分[18]。经非靶标代谢组学UHPLC-QTOF-MS分析,研究两种酵母培养物在正、负离子模式下,筛选出7 752和 6 709 个化合物,极大挖掘了更多代谢产物的信息,通过与数据库匹配,分别注释到4 940 和4 511 个化合物,两种酵母培养物进行了充分的发酵代谢,产生十分丰富的代谢产物,同时也包含大量未知代谢产物。其中,有机酸,核苷、核苷酸及其衍生物,氨基酸及其衍生物是占比较高的代谢物类别,这些丰富的营养物质,可刺激反刍动物瘤胃微生物生长,维持瘤胃厌氧环境,并提高发酵效率[19]。添加适量外源短链脂肪酸可改进瘤胃pH值和微生物种群,使有益微生物在瘤胃定植并加强瘤胃的发育,
参与瘤胃上皮细胞微生物屏障的构建,使之免受病
原微生物的侵害[20]。动物自身合成的核苷酸无法满足应激反应及快速生长的需要,酵母在各种微生物中所含核苷酸的比例最高,酵母培养物作为外源核苷酸的补充,可调节机体免疫力,促进幼龄动物生长[21]。此外,氨基酸可为反刍动物瘤胃中的乳酸利用菌提供营养物质,促进其增长繁殖,将过剩的乳酸转为短链脂肪酸,以维持pH值稳定的瘤胃环境[22]。
3.2
酵母培养物的代谢产物组成因不同生产商的菌种、培养基配方和发酵工艺等因素产生差异[23],是一类非标产品。在二级类别注释下,含量差异较大的代谢物可作为两种酵母培养物的特征代谢物。A组比B组相对含量高出16倍以上的差异代谢物有14种,其中染料木素、大豆苷元、白杨素和秦皮素是具药理作用的活性成分,有抗肿瘤、抗氧化、抑菌、调血脂、改善肠道菌群结构和组成等广泛药理活性[24-26]。同时,甘氨酸-L-亮氨酸可通过不同传递载体将靶向药物传递到所需器官或组织,在液体介质的生化过程中起重要作用[27]。此外,尿素可降低粗蛋白降解率,抑制有害菌群活动,提高牛羊消化率,增加反刍动物日增质量,从而提高青贮饲料的品质[28]。L-阿拉伯糖可调节血糖和血脂,改善代谢综合征,作为肠道细菌的碳源,可刺激有益菌繁殖,抑制致病菌增长,影响肠道微生物群落稳态及宿主健康[29]。另外,5-甲氧基色胺是合成褪黑激素的中间体,褪黑激素则具有节律平衡,改善睡眠、增强免疫力、清除自由基和延缓衰老等重要作用[30]。二氢辣椒素是激活瞬时受体电位香草型 1受体的激动剂,对心脏骤停后的缺血-再灌注脑损伤有保护作用[31]。1-甲基腺苷是甲基修饰的核苷酸,在生理条件下带有一个正电荷,其带来的电化学相互作用对于 tRNA 的功能和结构有着至关重要的作用[32]。B组比A组相对含量高出16倍以上差异代谢物有8种,其中,脱氢的硫酸雄甾酮可增强先天免疫系统,与其他肾上腺皮质系统调节对应激的适应性反应[33]。此外,4-羟基肉桂酸在氧化代谢应激条件下,能维持氧化还原稳态和能量代谢,具有神经保护作用[34]。而2-氧代己二酸是饲粮中必需氨基酸色氨酸和赖氨酸的关键代谢物,在尿液中的定量分析可反映氨基酸是否代谢异常而引发疾病[35]。另外,溶血磷脂酸是一种活跃的磷脂信号分子,可促进细胞增殖和分化,加强细胞间的相互作用,参与多种病理生理过程[36]。
3.3
天冬氨酸是合成甘氨酸、丝氨酸和苏氨酸的起始物质[37]。当碳源为糖蜜时,酵母在无氧条件下将其分解成半乳糖和葡萄糖混合物,随后优先快速利用优势碳源葡萄糖,而半乳糖作为非常规碳源,其代谢受葡萄糖抑制[38]。硫胺素即维生素B1,其功能形态焦磷酸硫胺素在细胞代谢中的行使多种辅助功能,米糠、麸皮、大豆及青绿牧草等饲料中含量较多,酵母本身也富含硫胺素[39]。生物素(Biotin)又名维生素H,是生物体内的一种辅酶,参与细胞多种代谢活动,可强烈促进酵母菌生长。玉米浆、麸皮水解液、甘蔗糖蜜等碳源原料都含有一定量的生物素,但不同生物素来源及其含量差异对微生物代谢有明显影响[40]。酵母培养物的生产通常在液体培养基进行有氧扩繁,随后在固态培养基进行厌氧发酵。糖代谢途径需经过中心物质丙酮酸,有氧时,通过柠檬酸循环彻底氧化,厌氧时,最终形成乙醇和CO2[37]。在柠檬酸循环中涉及柠檬酸、ɑ-酮戊二酸、琥珀酸、富马酸、L-苹果酸和草酰乙酸多种代谢物质,也是糖类、脂类、氨基酸代谢联系的枢纽。其中四碳二羧酸琥珀酸、富马酸、L-苹果酸和草酰乙酸因具有相似的化学结构可相互转化[41]。此外,酿酒酵母存在明显的Crabtree效应,即使氧气供应充足,酿酒酵母仍然会积累乙醇,这对柠檬酸循环十分不利,且基础碳源浓度越高,Crabtree效应越强[42]。
4" 结 论
在二级类别注释下,正、负离子模式分别注释到614、497个化合物,共237个差异代谢物在正离子模式检出,两种酵母培养物因碳氮源、发酵条件等不同因素,代谢产物含量存在显著差异。与达农威益康XP相比,自研酵母培养物发酵原料及工艺更简便。
参考文献(References)
[1]
Bu X Y, Lian X Q, Wang Y, et al. Dietary yeast culture modulates immune response related to TLR2-MyD88-NF-kβ signaling pathway, antioxidant capability and disease resistance against Aeromonas hydrophila for Ussuri catfish (Pseudobagrus ussuriensis)[J]. Fish amp; Shellfish Immunology," 2019, 84: 711-718.
[2] 杨东吉, 张静静, 朱随亮, 等. 酿酒酵母培养物替代抗生素对断奶仔猪生长性能、养分消化率及盲肠微生物区系的影响[J]. 动物营养学报, 2020, 32(1): 138-147.
YANG Dongji, ZHANG Jingjing, ZHU Suiliang, et al. Effects of Saccharomyces cerevisiae culture instead of antibiotics on growth performance, nutrient digestibility and cecal microflora of weaned piglets[J]. Chinese Journal of Animal Nutrition, 2020, 32(1): 138-147.
[3] 潘韵, 白静文, 梁子豪, 等. 7种酿酒酵母培养物成分分析及对仔猪的应用效果[J]. 粮食与饲料工业, 2019, (1): 49-52.
PAN Yun, BAI Jingwen, LIANG Zihao, et al. Composition analysis of seven Saccharomyces cerevisiae cultures and their effects on piglets[J]. Cereal amp; Feed Industry," 2019, (1): 49-52.
[4] 邵伟, 王立文, 张晓雪, 等. “益康XP” 对新疆荷斯坦奶牛生产性能和血清生化激素水平的影响[J]. 安徽农业科学, 2015, 43(21): 151-153.
SHAO Wei, WANG Liwen, ZHANG Xiaoxue, et al. Effects of“yikang XP” on production performance and serum biochemical hormone levels of Xinjiang Holstein cows[J]. Journal of Anhui Agricultural Sciences," 2015, 43(21): 151-153.
[5] 乔建国, 陈度煌, 余端梨, 等. 酵母培养物益康 “xp” 对母猪繁殖性能的影响[J]. 江西农业大学学报, 2007, 29(2): 266-269.
QIAO Jianguo, CHEN Duhuang, YU Duanli, et al. Effect of yeast culture yikang “xp” on reproductive performance of sows[J]. Acta Agriculturae Universitatis Jiangxiensis, 2007, 29(2): 266-269.
[6] 杨俊, 李毅, 许静, 等. 益康XP对肉种鸡产蛋性能及蛋品质的影响[J]. 广西畜牧兽医, 2013, 29(3): 131-132.
YANG Jun, LI Yi, XU Jing, et al. Effects of yikang XP on egg performance and egg quality of broilers[J]. Guangxi Journal of Animal Husbandry amp; Veterinary Medicine, 2013, 29(3): 131-132.
[7] 杨越. 益康XP替代抗生素对断奶仔猪生长性能和血清生化指标的影响[J]. 黑龙江畜牧兽医, 2014,(5): 85-87.
YANG Yue. Effects of Yikang XP replacing antibiotics on growth performance and serum biochemical indexes of weaned piglets[J]. Heilongjiang Animal Science and Veterinary Medicine, 2014,(5): 85-87.
[8] 吕凯, 侯生珍, 郭远玉, 等. “益康XP” 替代抗生素对早期断奶藏羔羊生长发育的影响[J]. 中国农学通报, 2012, 28(32): 21-25.
LYU Kai, HOU Shengzhen, GUO Yuanyu, et al. Effects of diamond V “XP” -yeast culture instead of antibiotics on growth and development in early-weaned lambs[J]. Chinese Agricultural Science Bulletin, 2012, 28(32): 21-25.
[9] 孙喆. 酵母培养物有效物组的研究[D]. 长春: 吉林农业大学, 2018: 30-96.
SUN Zhe. Study on the Effective Compounds Group of Yeast Culture[D]. Changchun: Jilin Agricultural University, 2018: 30-96.
[10] 郭永清, 赵宇飞, 张小宇. 酵母培养物对断奶犊牛生长性能及瘤胃发酵的影响[J]. 饲料研究, 2019, 42(11): 10-13.
GUO Yongqing, ZHAO Yufei, ZHANG Xiaoyu. Effect of yeast culture on growth performance and rumen fermentation of weaned calves[J]. Feed Research, 2019, 42(11): 10-13.
[11] 詹世琛, 马建宁. 酵母培养物在奶牛生产中的应用概况及发展前景[J]. 中国奶牛, 2015,(17): 1-6.
ZHAN Shichen, MA Jianning. Application and development prospect of yeast culture in dairy cow production[J]. China Dairy Cattle," 2015,(17): 1-6.
[12] Fiehn O, Kopka J, Drmann P, et al. Metabolite profiling for plant functional genomics[J]. Nature Biotechnology, 2000, 18(11): 1157-1161.
[13] Sumner L W, Mendes P, Dixon R A. Plant metabolomics: large-scale phytochemistry in the functional genomics era[J]. Phytochemistry, 2003, 62(6): 817-836.
[14] Fiehn O. Metabolomics—the link between genotypes and phenotypes[J]. Plant Molecular Biology, 2002, 48(1/2): 155-171.
[15] 鲁晓宇, 郭家鑫, 陶一凡, 等. 不同盐碱胁迫对棉花营养元素吸收转运以及代谢的影响[J]. 中国生态农业学报(中英文), 2023, 31(3): 438-451.
LU Xiaoyu, GUO Jiaxin, TAO Yifan, et al. Effects of different salt and alkali stress on absorption, transportation, and meta-bolism of nutrient elements in cotton[J]. Chinese Journal of Eco-Agriculture, 2023, 31(3): 438-451.
[16] Berrueta L A, Alonso-Salces R M, Héberger K. Supervised pattern recognition in food analysis[J]. Journal of Chromatography A, 2007, 1158(1/2): 196-214.
[17] 张得香, 朱超, 周加义, 等. 酵母培养物促进干细胞增殖分化、改善小鼠肠上皮完整性[J]. 动物营养学报, 2022, 34(6): 3962-3971.
ZHANG Dexiang, ZHU Chao, ZHOU Jiayi, et al. Yeast culture promotes stem cell proliferation and differentiation and improves intestinal epithelial integrity of mice[J]. Chinese Journal of Animal Nutrition, 2022, 34(6): 3962-3971.
[18] 彭一凡, 甄玉国. 酵母培养物及其在养殖业中的应用[J]. 饲料工业, 2008, 29(10): 30-33.
PENG Yifan, ZHEN Yuguo. Application of yeast culture in livestock breeding[J]. Feed Industry, 2008, 29(10): 30-33.
[19] McAllister T A, Beauchemin K A, Alazzeh A Y, et al. Review: the use of direct fed microbials to mitigate pathogens and enhance production in cattle[J]. Canadian Journal of Animal Science, 2011, 91(2): 193-211.
[20] 张剑霞, 胡红莲, 宋利文, 等. 短链脂肪酸对亚急性瘤胃酸中毒的影响和钠离子耦合单羧酸转运蛋白1和氢离子耦合单羧酸转运蛋白1对短链脂肪酸的转运机制[J]. 动物营养学报, 2022, 34(12): 7574-7584.
ZHANG Jianxia, HU Honglian, SONG Liwen, et al. Effects of short chain fatty acids on subacute ruminal acidosis and transport mechanism of sodium-coupled monocarboxylate transporter 1 and hydrogen-coupled monocarboxylate transporter 1 on short chain fatty acids[J]. Chinese Journal of Animal Nutrition, 2022, 34(12): 7574-7584.
[21] 王会. 日粮添加酵母核苷酸对断奶仔猪生长性能、血清生化指标及免疫力的影响[J]. 饲料研究, 2021, (18): 36-39.
WANG Hui. Effect of dietary supplementation of yeast nucleotides on growth performance, serum biochemical indexes and immunity of weaned piglets[J]. Feed Research, 2021, (18): 36-39.
[22] Dias A L G, Freitas J A, Micai B, et al. Effects of supplementing yeast culture to diets differing in starch content on performance and feeding behavior of dairy cows[J]. Journal of Dairy Science, 2018, 101(1): 186-200.
[23] 张弦, 章亭洲, 瞿明仁. 活性干酵母及酵母培养物在反刍动物中的研究进展与应用[J]. 动物营养学报, 2022, 34 (1): 20-29 .
ZHANG Xian, ZHANG Tingzhou, QU Mingren. Research progress and application of active dry yeast and yeast culture in ruminants [J]. Chinese Journal of Animal Nutrition, 2022, 34(1): 20-29.
[24] 高葱葱, 葛稳, 方怡超, 等. 基于16S rRNA测序技术研究染料木素和大豆苷元对大鼠肠道菌群的影响[J]. 中草药, 2023, 54(3): 834-840.
GAO Congcong, GE Wen, FANG Yichao, et al. Effect of genistein and daidzein on gut microbiota in rats based on 16S rRNA sequencing[J]. Chinese Traditional and Herbal Drugs, 2023, 54(3): 834-840.
[25] 凡春玲, 申栋帅, 冯小明, 等. 白杨素衍生物的抗氧化和抗炎活性研究进展[J]. 中国兽药杂志, 2022, 56(11): 69-78.
FAN Chunling, SHEN Dongshuai, FENG Xiaoming, et al. The review of antioxidant and anti-inflammatory activity of chrysin derivatives[J]. Chinese Journal of Veterinary Drug, 2022, 56(11): 69-78.
[26] 李澎瀛, 李京, 何金环, 等. 秦皮素的生物活性及药物作用机制研究进展[J]. 现代牧业, 2021, 5(4): 51-56.
LI Pengying, LI Jing, HE Jinhuan, et al. Research progress on the biological activities and drug mechanisms of fraxetin[J]. Modern Animal Husbandry, 2021, 5(4): 51-56.
[27] Lytkin A I, Chernikov V V, Krutova O N, et al. Thermodynamics of the dissolution of glycyl-L-leucine in aqueous solutions[J]. Russian Journal of Physical Chemistry A, 2020, 94(1): 77-80.
[28] 谷丽, 杨磊, 王祎. 青贮饲料添加剂的研究进展[J]. 农业技术与装备, 2022,(8): 87-89.
GU Li, YANG Lei, WANG Yi. Research progress of silage additives[J]. Agricultural Technology amp; Equipment, 2022,(8): 87-89.
[29] 倪娓娓, 胡超群, 白福来. L-阿拉伯糖的生理功能及市场应用进展[J]. 中国食品添加剂, 2022, 33(1): 16-20.
NI Weiwei, HU Chaoqun, BAI Fulai. Physiological function and application progress of L-arabinose[J]. China Food Additives, 2022, 33(1): 16-20.
[30] 胡松林, 蔡瑾, 须辑. N-乙酰基-5-甲氧基色胺的化学合成[J]. 化学世界, 2016, 57(11): 699-702.
HU Songlin, CAI Jin, XU Ji. Chemical synthesis of melatonin[J]. Chemical World, 2016, 57(11): 699-702.
[31] 钟晓芃, 郭义, 张诗武. 二氢辣椒素不依赖诱导亚低温对心脏骤停复苏后脑损伤的防治作用及分子机制[J]. 中国急救医学, 2022, 42(10): 850-857.
ZHONG Xiaopeng, GUO Yi, ZHANG Shiwu. Protective effect and molecular mechanism of dihydrocapsaicin independent of induced mild hypothermia on brain injury after the resuscitation in cardiac arrest patients[J]. Chinese Journal of Critical Care Medicine, 2022, 42(10): 850-857.
[32] Agris P F. The importance of being modified: roles of modified nucleosides and Mg2+ in RNA structure and function[J]. Progress in nucleic acid research and molecular biology, 1996, 53: 79-129.
[33] Buhimschi C S, Turan O M, Funai E F, et al. 200: intra-amniotic inflammation (IAI) is associated with a low fetal plasma cortisol/dehydroepi- androsterone sulfate ratio (fetal stress index)[J]. American Journal of Obstetrics and Gynecology," 2007, 197(6): S67.
[34] Park S J, Kim Y A, Lee J W, et al. 4-Hydroxycinnamic acid attenuates neuronal cell death by inducing expression of plasma membrane redox enzymes and improving mitochondrial functions[J]. Food Science and Human Wellness, 2023, 12(4): 1287-1299.
[35] Shibata K, Yasui M, Sano M, et al. Fluorometric determination of 2-oxoadipic acid, a common metabolite of tryptophan and lysine, by high-performance liquid chromatography with pre-chemical derivatization[J]. Bioscience, Biotechnology, and Biochemistry, 2011, 75(1): 185-187.
[36] 张燕燕, 王靖雷, 潘阳阳, 等. 溶血磷脂酸受体1-3在牦牛不同时期卵巢中表达定位[J]. 动物医学进展, 2021, 42(9): 35-42.
ZHANG Yanyan, WANG Jinglei, PAN Yangyang, et al. Lysophosphatidic acid receptors LPAR1-3Expressions and localizations in yak ovaries at different stages[J]. Progress in Veterinary Medicine, 2021, 42(9): 35-42.
[37] 龚大春, 彭艳, 涂璇, 等. 酵母生物学[M]. 北京: 中国水利水电出版社, 2015: 77-78.
GONG Dachun, PENG Yan, TU Xuan, et al. Yeast biology[M]. Beijing: China Water amp; Power Press, 2015: 77-78.
[38] 邹静, 孟军, 张建才, 等. 利用基因工程提高酿酒酵母半乳糖代谢[J]. 河北科技师范学院学报, 2016, 30(4): 14-22.
ZOU Jing, MENG Jun, ZHANG Jiancai, et al. Improvement of galactose metabolism of Saccharomyces cerevisiae via gene engineering[J]. Journal of Hebei Normal University of Science amp; Technology, 2016, 30(4): 14-22.
[39] 姚婉婷, 李可, 宋娜, 等. 硫胺素对鲁氏接合酵母高盐适应性的影响[J]. 中国酿造, 2020, 39(11): 78-84.
YAO Wanting, LI Ke, SONG Na, et al. Effect of thiamine on high-salt adaptability of Zygosaccharomyces rouxii[J]. China Brewing, 2020, 39(11): 78-84.
[40] 黄瑶, 廖兰, 廖春燕, 等. 不同来源生物素对谷氨酸短杆菌的影响[J]. 广西工学院学报(自然科学版), 2010, 21(3): 80-83.
HUANG Yao, LIAO Lan, LIAO Chunyan, et al. Effects of biotin from several materials on Brevibacterium glutamicum[J]. Journal of Guangxi University of Technology (Natural Science), 2010, 21(3): 80-83.
[41] Cao X F, Yan L Q, Han L X, et al. Association of mild to moderate kidney dysfunction with coronary artery calcification in patients with suspected coronary artery disease[J]. Cardiology, 2012, 120(4): 211-216.
[42] 刘立聪, 汤超, 王志, 等. 甘油和生物素对酿酒酵母产胞内谷氨酸的影响[J]. 湖北工业大学学报, 2017, 32(2): 42-46.
LIU Licong, TANG Chao, WANG Zhi, et al. Effects of Glycerol and Biotin additions on intracellular Glutamate Production by Saccharomyces cerevisiae[J]. Journal of Hubei University of Technology, 2017, 32(2): 42-46.
Analysis of the composition differences between two yeast cultures based on untargeted metabolomics
LIN Qing1, SHI Hongling1, QIN Xinzheng1, LI Yue1, WANG Zihan1, GAO Yan1, ZENG Jun1, WANG Haozhong2, LOU Kai1, HUO Xiangdong1
(1. Xinjiang Laboratory of Special Environmental Microbiology / Institute of Applied Microbiology, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China;2.College of Life Science and Technology, Xinjiang University, Urumqi 830046, China )
Abstract:【Objective】 This project aims to compare the metabolic components differences between the XP and experimental group yeast culture,the findings will provide a reference for guiding the production and application of yeast culture.
【Methods】 The untargeted metabolomics UHPLC-QTOF-MS technology was used to analyze and compare the composition and differences of the metabolic components between the 2 yeast cultures.
【Results】" (1) Under the super classification annotation, a total of 614 and 497 compounds were noted in positive and negative ion modes respectively.The main metabolic classes were organic acids, nucleosides, nucleotides and their derivatives, amino acids and their derivatives and so on.There were no specific metabolic components in the two yeast cultures, only significant differences in content (Plt;0.05).(2) In the positive ion mode, 237 differential metabolites were detected, of which 176 were significantly up-regulated(Plt;0.05) and 61 were significantly down-regulated (Plt;0.05) in expression.In the negative ion mode, 136 differential metabolites were detected, of which 64 were significantly up-regulated (Plt;0.05) and 72 were significantly down-regulated (Plt;0.05) in expression.(3) The KEGG (Kyoto Encyclopedia of Genes and Genomes) enrichment pathway of differential metabolites mainly focused on alanine, aspartate and glutamate metabolism.
【Conclusion】" Due to different carbon and nitrogen sources and fermentation conditions, the yeast cultures have significant differences in the content of metabolites, but with their own advantages of the biological activity of main differential metabolites.
Key words:yeast culture; untargeted metabolomics; differential metabolite; Saccharomyces cerevisiae
Fund projects:Key Program of Natural Science Foundation of" Xinjiang Uygur Autonomous Region (2023D01D11);
Key program of scientific and technological research plan of Xinjiang production and construction corps(2022AB013);Key program of scientific and technological research plan of Xinjiang production and construction corps(2020AB013)
Correspondence author: LOU Kai(1968-),male,from Henan, researcher, master tutor, Ph.D.,research direction: microbial ecology,(E-mail)@tsinghua.org.cn
HUO Xiangdong (1974-), male, from Gansu, associate professor, Ph.D., research direction:microbial resources,(E-mail) xiangdonghuo@163.com