摘要: "【目的】种源与家系早期选择是林木育种的重要措施,通过分析不同地理种源观光木的表型性状,以期揭示观光木表型变异程度及变异规律,为观光木优良种质资源的优良选种、保护和开发利用提供理论基础。【方法】本研究以前期收集的11个地理种源1年生实生苗为对象,对不同地理种源观光木苗期生长节律进行初步分析;比较表型性状在种源间的差异和变异系数在种源间的变化以及与种源地理、气候因子的关系,并分析观光木幼苗各器官生物量在种源间的差异,采用聚类分析和模糊隶属函数进行种源综合评价。【结果】观光木在苗期生长过程均呈现明显的“慢—快—慢”的“S”型生长节律,生长速生期在7~10月;不同地理种源观光木表型性状指标、生物量均有显著差异(P<0.01),南宁树木园种源的表型性状指标在11个种源中均处于优势;不同地理种源表型性状平均变异系数为14.11%,苗高、地径的差异明显大于其他指标的差异;从生物量的变异系数上来看,观光木各种源间茎鲜重的变异系数要大于根鲜重,但整体上看,茎鲜重也高于根鲜重,说明观光木各种源间的生物量具有一定的稳定性。绝大部分苗期表型性状与地理因子呈显著负相关关系,观光木在低纬度、温度高、降水充沛的地方生长良好;聚类分析结果和模糊隶属函数评价结果具有一致性,将11个观光木种源划分为3类,初步筛选南宁树木园种源为优良种源。【结论】观光木11个种源苗期表型性状和生物量变异丰富,说明观光木优良种源在苗期就有较大的选择潜力,能够进行基于表型性状的种质资源筛选。
关键词: "观光木; "种源; "苗期; "表型性状; "生物量
中图分类号: " S 567. 19 " " " " " " " 文献标识码: " A " " " " " " " "文章编号:1001 - 9499(2024)04 - 0001 - 07
Phenotypic Traits and Biomass Comparative Analysis of Michelia odora Seedlings from Different Provenances
ZHOU Fan1 WANG Yiping1** XIONG Yang1 WU Zhaoxiang2 YANG Zhijun1
MEI Yufeng3 DU Jian3 YUAN Wenbin4
(1. "Gannan Arboretum, "Jiangxi Ganzhou 341212; "2. "Institute of Biological Resources, Jiangxi Academy of Sciences, "Jiangxi Nanchang 330096; "3. "Ganzhou Forestry Ecological Service Center, "Jiangxi Ganzhou 341000; "4. "Gold Rooster Forest Farm of Xinfeng County, "Jiangxi Ganzhou 341600)
Abstract For the early selection of superior provenance for Michelia odora, based on the one-year seedlings of 11 geographical provenances collected previously, the differences in phenotypic traits, the changes in coefficient of variation among provenances, as well as their relationships with geographical and climatic factors were compared, and the differences in biomass of various organs of M. odora seedlings among provenances were analyzed. Taking advantage of the method of cluster analysis and fuzzy membership function, the comprehensive evaluation of the M. odora provenance was carried out. The results showed that the growth increment of "M. odora during the seedling stage exhibited significant rhythmicity. The growth rate decreased significantly from July to October, and after November, it reached the lowest point and almost stopped growing. Phenotypic traits and biomass differed significantly among geographical provenances of M. odora (Plt;0.01). The average coefficient of variation of phenotypic traits from different geographical provenance was 14.11%, and the majority of phenotypic traits at seedling stage correlated negatively with geographical factors significantly. M. odora grew very well in low latitude, high temperature and abundant precipitation areas. The 11 M. odora geographical provenances was divided into 3 groups by the clustering analysis and the fuzzy membership function evaluation, and the M. odora from Nanning Arboretum was identified as the superior one.
Key words Michelia odora; seedling stage; provenance; phenotypic trait; biomass
种源或地理种源是获得种子或繁殖材料的原产地。种源试验是将地理种源不同的种子或其他繁殖材料放到一起所做的栽培对比试验[ 1 ]。同一个树种在自然分布区内,由于种源的不同,受各地气候、土壤及经营条件的影响也不同,就形成了各种不同的生态型,在苗木性状及林分生态上产生差异[ 2 ]。幼苗生长性状和生物量情况是最能直接反映苗木生长状况的指标[ 3 ]。幼苗生长性状和生物量情况分析主要通过方差分析、相关性分析、遗传参数估计、多重比较和聚类分析等统计方法得到结果[ 4 ]。因林木生长周期长,在筛选优良种源的过程中,通常选择早期选择的方法。我国通过早期选择对桢楠(Phoebe zhennan)、火力楠(Michelia macclurei)、油松(Pinus tabuliformis)、枫香(Liquidambar formosana)、马褂木(Liriodendron chinense)、米老排(Mytilaria laosen- "sis)、核桃楸(Juglans mandshurica)、大叶栎(Quercus griffithii)、樟(Camphora officinarum)、杉木(Cunning- "hamia lanceolata)、云南松(Pinus yunnanensis)等树种[ 5 - 15 ]进行种源评价及优良种源选择。
观光木(M. odora)为木兰科(Magnoliaceae)含笑属(Michelia)的常绿树种,又叫香花木、香木楠、宿轴木兰,是中国特有的古老孑遗树种,其零散分布于江西南部、福建、广东、海南、广西、云南东南部等海拔300~1100 m的岩山地常绿阔叶林中。由于生态环境恶化、人类干扰及盗伐现象严重,观光木野生资源数量急剧减少,加之自然更新能力较差、种群天然分布狭窄,1984年被列入国家珍稀濒危二级保护植物。观光木是中国中部和南方特有的著名园林绿化和香料植物,它是继同科植物乐昌含笑(M. chapensis)、深山含笑(M. maudiae)、红花木莲(Manglietia insignis)之后的又一新树种。观光木是木兰科中较进化种类,对木兰科的分类系统研究有重要意义。此外,观光木对研究古代植物区系、古地理、古气候都有着重要的科学价值。
近年来国内观光木的研究方向主要集中在观光木的生物量、引种繁殖、群落与种群生态、系统发育等基础生物学方面。关于苗期生长规律研究仅有池毓章[ 16 ]于2007 年研究过观光木播种育苗生长规律及育苗技术,对其生长过程、种子贮藏与播种育苗管理技术有较详细记述。程世[ 17 ]等研究观光木播种苗的年生长规律,表明从种子发芽出土至6月期间生长慢,7月生长速度中等,8~10月期间生长最快,11~12月生长较慢甚至停止生长。吉悦娜[ 18 ] 采用大田试验方法,揭示了冬播与春播对观光木场圃发芽率差异不大,分别为56%和55%,但冬播能提高苗木的生长量和苗木质量。然而,从表型性状以及生物量的角度分析不同地理种源观光木苗期生长变异状况的研究还未见报道。本研究基于前期收集的4省11县共11个观光木种源材料,以1年生实生苗为研究对象,通过对表型性状和生物量的测定,比较表型性状和生物量在种源间的差异;同时结合种源地理因子分析各因子对观光木子代苗期表型性状的影响,旨在了解不同种源观光木苗期生长性状,初步筛选出在苗期生长表现较好的观光木优良种源,为观光木人工林培育中进行种源选择提供参考。
1 试验地概况
赣南树木园位于罗霄山脉东侧,园区坐落在阳明湖国家森林公园内。处于上犹江中游,地跨上犹、崇义两县。地理位置为114°02~114°04E,25°50~25°51N,海拔198~633 m。东南面被湖水环绕,西北面以高山岭脊为界。整个园区由十二个半岛和岛屿组成,属中亚热带季风区山地湿润气候类型,四季分明,年均温17.8 ℃,年降水量1 615.2 mm,年蒸发量1 031.5 mm,无霜期302天,相对湿度80%以上。土壤以黄红壤为主,pH值5.0~5.5,非常适宜壳斗科、樟科、冬青科等多种亚热带常绿阔叶树种以及马尾松、杉木等一批用材树种的生长和发育。
2 材料与方法
2. 1 试验材料
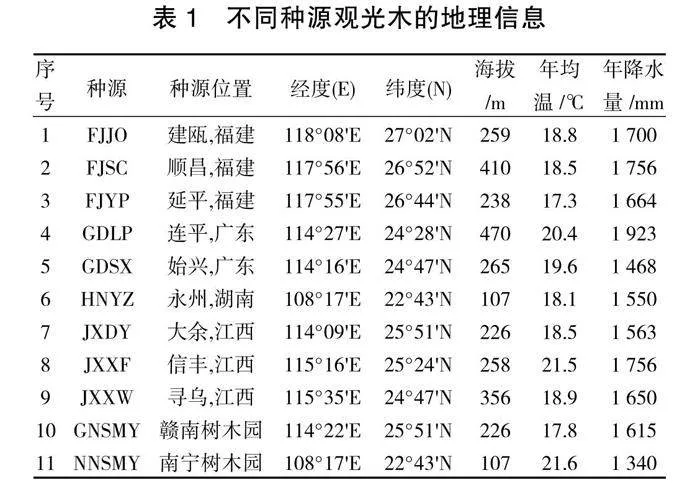
试验材料来自4省11县市的11个地理种源(表1),于2021年10月采集11个种源地成熟种子,经湿沙储藏后,2022年3月在园区中洞苗圃基地以条播的方式进行播种育苗,设置行距为20 cm,深度为3~5 cm,将种子置于播种沟内并覆盖1~1.5 cm厚细土,播种后覆盖5 cm厚先前准备好的铁芒萁枝条。播种后20~30天左右开始出苗,出苗整齐后揭开铁芒萁。在6月份进行间苗补苗,每行保留苗木10~15株。在试验期间,除草、水肥、病虫害防治等苗期管理均按照常规方法进行。
2. 2 苗高、地径生长指标测定
自2022 年7月份开始测量观光木第一年的苗高、地径。每个种源随机选择30株苗木,挂牌标记,作为苗木生长量测定的固定标准株,2个月测量1次,并在2023年2月结合生物量数据收集进行1次年终生长量测定。苗高用钢尺测量(精确到0.1 cm),地径用游标卡尺测量(精确到0.1 mm)。
2. 3 苗期性状和生物量的测定
2023年2月对幼苗进行表型性状和生物量的测定与获取。每个种源选取30株生长健康且无病虫害的苗木作为标准株。取苗前对叶片数进行准确计数,然后将观光木幼苗从苗圃连根挖起,将根系泥土清洗干净。用枝剪将植株分为根、茎、叶三种不同植物器官,分单株测量苗高、地径等生长指标,根、茎的鲜质量、主根长和叶片数量等生物量指标。
(1)表型性状测定
每个标准株随机选取3片新鲜老叶测定叶片性状,采用叶面积测定仪(YMJ-A)获取叶长(cm)、叶宽(cm)、叶面积(cm2),叶绿素(SPAD)、碳含量(mg/g)用叶绿素测定仪(北京中科维禾TYS-4N)测量。
(2)生物量测定
分单株测量标准株根、茎的鲜质量,主根长和叶片数量,用电子天平测定鲜重(精确度0.01 g),主根长用钢尺测量(精确到0.1 cm),根据根茎鲜重计算出茎根比:茎根比=茎鲜重/根鲜重。
2. 4 数据分析方法
用Excel 2019软件进行基础数据的整理、统计,运用SPSS 25.0软件进行统计分析。通过方差分析比较不同种源观光木苗期各表型性状、生物量,同时,计算表型性状与种源地理、气候因子之间的Spearman相关系数,采用欧氏距离进行聚类分析。采用模糊数学隶属函数法对不同地理种源观光木进行综合评价。
2. 5 模糊数学隶属函数法
采用模糊数学隶属函数法[ 19 ]对观光木的表型性状进行综合评价。隶属函数值X(μ)=(X - Xmin)/( Xmax - Xmin),式中x为指标测定值;Xmin、Xmax为所有参试材料某一指标的最小值和最大值。如果某一指标与表型性状呈负关,则可以通过反隶属函数计算其隶属函数值,公式为X(v)=1 -(X-Xmin)/(Xmax-Xmin)。计算隶属函数值平均值并作为优良种源评价指标,值越大,表明该种源的优良度越高。
3 结果与分析
3. 1 不同地理种源观光木苗期生长节律初步分析
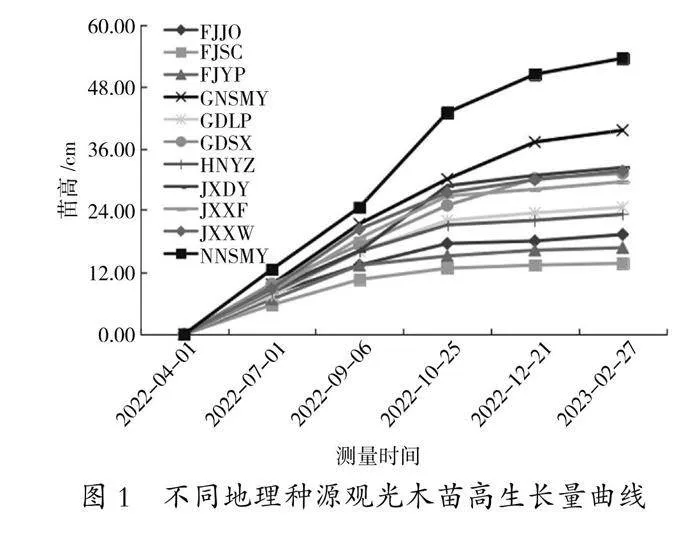
由图1和图2可知,11个种源观光木在4月整体出苗至7月开始数据观测期间生长较为缓慢,7月至10月下旬苗高生长速度较快,11月之后苗高生长缓慢。苗高快速生长的时间持续约4个月。观光木苗高年生长过程均呈现明显的“慢—快—慢”的“S”型生长节律。且苗高在整个生长季节都持续增长,属于全期生长类型。对不同种源观光木进行比较发现,9 月上旬以前各个种源的苗高生长差异很小;从9 月初开始到10 月下旬,不同种源间高生长差异逐渐拉大,生长最快的南宁树木园(NNSMY)和生长最慢的福建顺昌(FJSC)种源相差30.18 cm,前者约为后者的4 倍。此后,虽然赣南树木园(GNSMY)高增长略快于其他种源,但是各种源的生长差异趋于稳定。
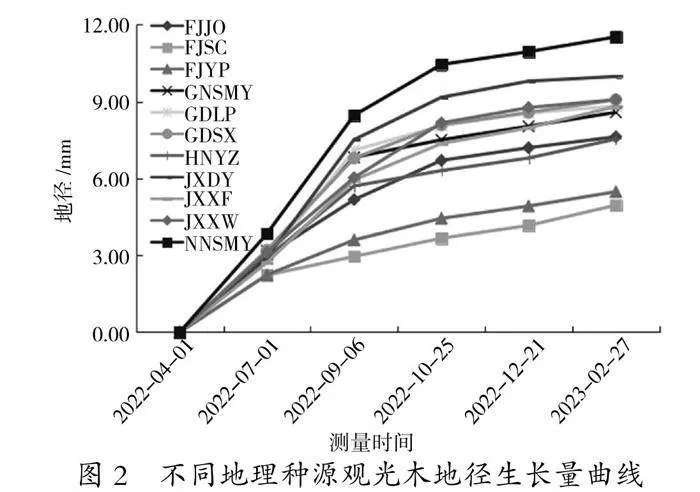
从观光木各种源地径生长过程(图2)可知,各种源地径生长趋势比较一致,但依旧表现出“慢—快—慢”的趋势。各种源间地径的生长差异在9月6日之后逐渐拉大,至生长停止时福建延平(FJYP)、福建顺昌(FJSC)地径显著小于其他种源,比最粗的南宁树木园(NNSMY)小6.04~6.57 mm,前者仅为后者的43%左右。除南宁树木园(NNSMY)、江西大余(JXDY)、福建延平(FJYP)、福建顺昌(FJSC)种源外,其余7个种源间地径差异较小。
3. 2 不同地理种源观光木苗期表型性状差异
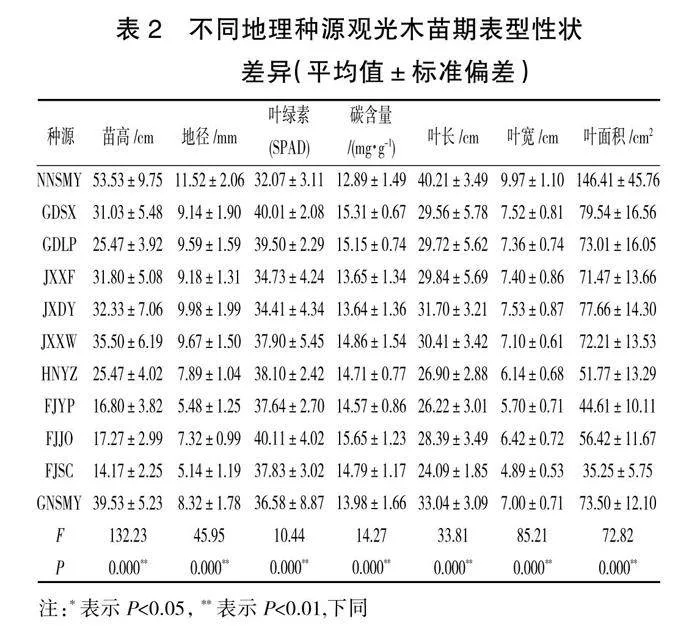
观光木苗期表型性状在不同种源间的差异性见表2,经F检验,观光木11个种源7个表型性状差异极显著(P<0.01),其中苗高、地径、叶面积差异最明显。苗高、地径、叶长、叶宽、叶面积等指标在南宁树木园(NNSMY)表现最好,而在福建顺昌(FJSC)表现最差,南宁树木园(NNSMY)各个指标分别达到福建顺昌(FJSC)的3.78、2.24、1.67、2.04、4.15倍。叶绿素和碳含量在福建建瓯(FJJO)表现最好,在南宁树木园(NNSMY)表现最差。南宁树木园(NNSMY)的苗高、地径、叶长、叶宽、叶面积等指标在11个种源中均处于优势,初步筛选南宁树木园(NNSMY)为优良速生种源。
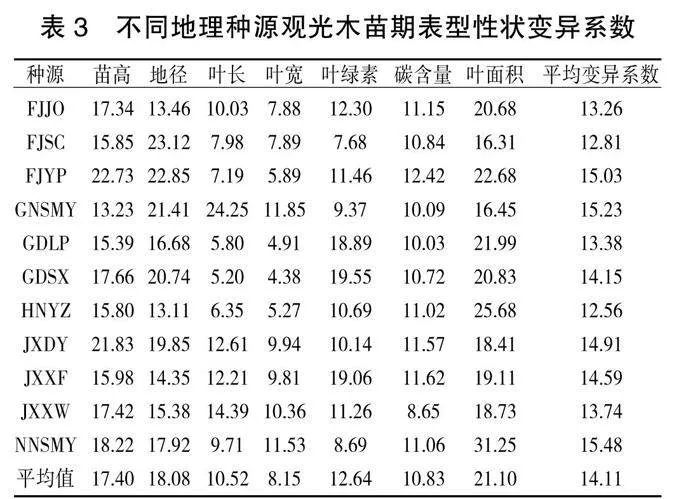
3. 3 不同地理种源观光木苗期表型性状的形态变异特征
由表3可知,观光木各性状平均变异系数为14.11%,变异幅度为8.15%~21.10%,最大变异性状变异系数(苗高)是最小变异系数(叶宽)的2.6倍,7个表型性状的平均变异系数有一定的差异,其平均变异系数为17.4%、18.08%、10.52%、8.15%、12.64%、10.83%和21.10%。苗高、地径、叶面积的变异系数在20%左右,叶长、叶宽、叶绿素、碳含量的变异系数在10%左右,表明苗高、地径、叶面积的差异性高于叶长、叶宽、叶绿素、碳含量。11个种源所有性状的平均变异系数为12.56%~15.48%,南宁树木园(NNSMY)的平均变异系数最大(15.48%),湖南永州(HNYZ)的平均变异系数最小(12.56%),表明湖南永州(HNYZ)的表型性状稳定性较高。从变异系数来看,不同种源间观光木苗高、地径的差异明显大于其他指标的差异。
3. 4 不同地理种源观光木苗期表型性状与种源地理的相关性
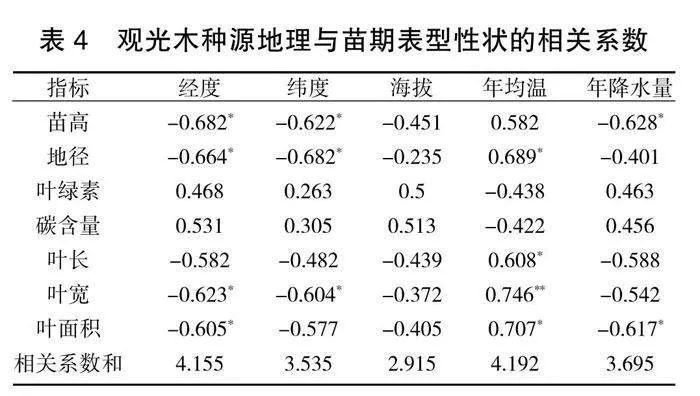
对不同种源观光木的表型性状指标与地理因子、气候因子进行相关分析与检验,比较地理、气候因子对表型性状的综合相关性。结果(表4)表明,相关性系数总和大小依次为:年均温(4.192)>经度(4.155)>年降水量(3.695)>纬度(3.535)>海拔(2.915)。不同种源观光木苗高、地径、叶宽、叶面积与经度、纬度呈显著负相关关系(P<0.05);海拔与其他指标之间均不会呈现出显著性;年均温与叶宽呈极显著正相关关系(P<0.01);年降水量与苗高、叶面积呈显著负相关关系(P<0.05)。地理因子与观光木苗期各表型性状的相关性绝大部分为显著负相关,表明地理因子对1年生观光木幼苗表型性状的影响存在一定的差异性。说明观光木在低纬度、温度高、降水充沛的地方生长良好。
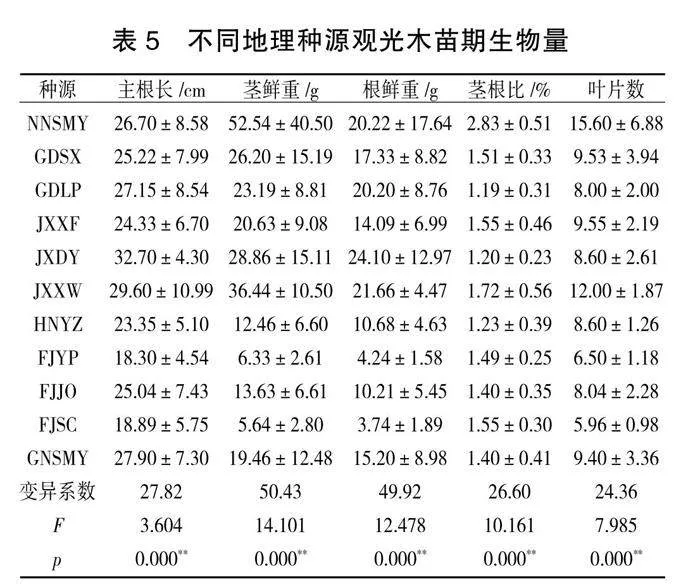
3. 5 不同地理种源观光木苗期生物量差异
利用方差分析去研究种源对于主根长、茎鲜重、根鲜重、茎根比、叶片数共5项的差异性。由表5可知,不同种源样本的主根长、茎鲜重、根鲜重、茎根比、叶片数全部呈现出极显著性差异(P<0.01)。不同种源间生物量的变异系数集中在20%~50%。不同种源生物量种内变异十分明显,其中变异程度从大到小为茎鲜重gt;根鲜重gt;主根长gt;茎根比gt;叶片数。江西大余(JXDY)的主根长和根鲜重最大,显著高于其他种源,分别达到(32.70±4.30)cm、(24.10± "12.97)g,说明江西大余(JXDY)的根系比其他种源的发达;不同种源间地上部分茎鲜重和叶片数以南宁种源(NNSMY)和江西寻乌(JXXW)的较高,福建顺昌(FJSC)和福建延平(FJYP)较低,茎鲜重最大值南宁种源(NNSMY)是最小值福建顺昌(FJSC)的9.31倍,南宁种源(NNSMY)和江西寻乌(JXXW)的叶片数在10片以上,其他种源均在10片以下。茎根比是苗木地上部分与地下部分(重量或体积)之比,反映出苗木根茎两部分的平衡状况[ 21 ],茎根比越大表明地上部分发达而根系弱,茎根比小说明根系发达,苗木健壮,茎根比以南宁种源(NNSMY)显著高于其他种源,是最小值广东连平(GDLP)的2.38倍。从变异系数上来看,观光木各种源间茎鲜重的变异系数要大于根鲜重,但整体上看,茎鲜重也高于根鲜重。说明观光木各种源间的生物量具有一定的稳定性。
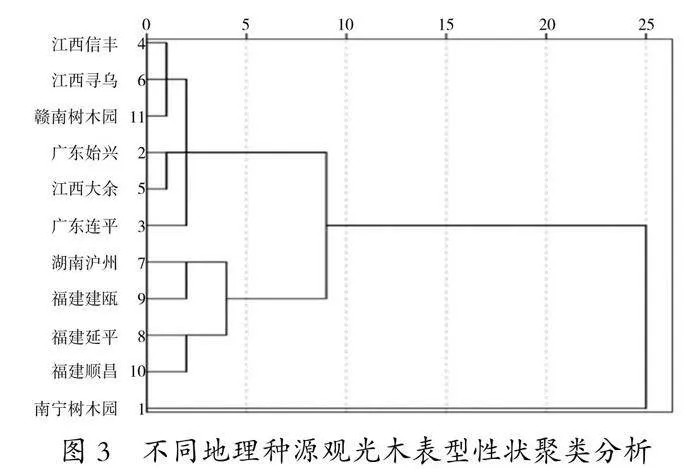
3. 6 不同地理种源观光木表型性状的聚类分析
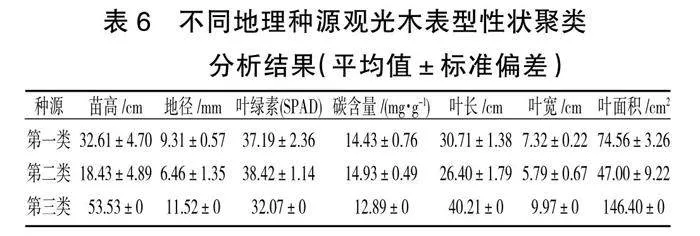
对11个观光木种源苗高、地径、叶绿素、碳含量、叶长、叶宽、叶面积等指标进行系统聚类分析(图1),当欧氏距离D=9.0时,将11个观光木种源分为3个类群,分别用A、B、C表示。
A类含6个种源,占比54.55%,包括江西信丰(JXXF)、江西寻乌(JXXW)、赣南树木园(GNSMY)、广东始兴(GDSX)、江西大余(JXDY)、广东连平(GDLP);A类为一般种源,该类群的指标均处于中等水平。
B类含4个种源,占比36.36%,包括湖南永州(HNYZ)、福建建瓯(FJJO)、福建延平(FJYP)、福建顺昌(FJSC);该类种源的叶绿素、碳含量平均值为最高,分别为(38.42±1.14)、(14.93±0.49)mg/g;苗高、地径、叶长、叶宽、叶面积平均值均为最低。
C类含1个种源,占比9.09%,为南宁种源(NNSMY),该种源的苗高、地径、叶长、叶宽、叶面积平均值均最大,分别为53.53 cm、11.52 cm、40.21 cm、9.97 cm、146.40 cm2。
3. 7 观光木的模糊综合评价
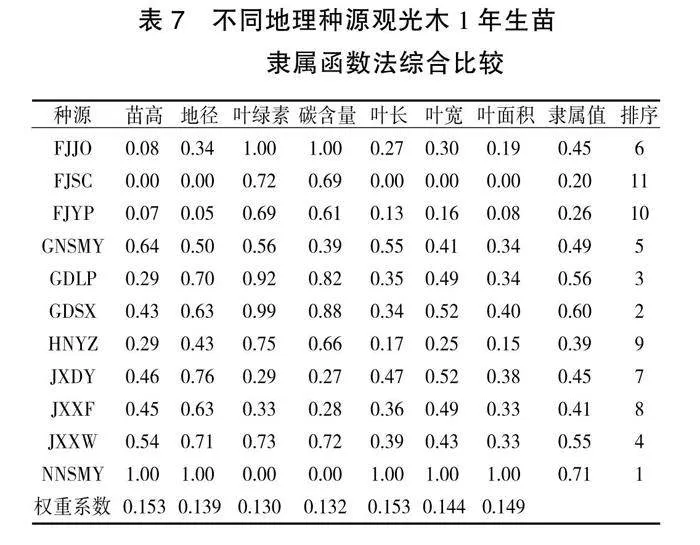
采用模糊隶属函数对观光木的苗高、地径、叶绿素、碳含量、叶长、叶宽、叶面积等7个表型性状指标进行综合评价,判断不同地理种源观光木幼苗的优良度(表7)。不同地理种源观光木表型性状的隶属函数值大小依次为:南宁树木园(NNSMY)>广东始兴(GDSX)>广东连平(GDLP)>江西寻乌(JXXW)>赣南树木园(GNSMY)>福建建瓯(FJJO)>江西大余(JXDY)>江西信丰(JXXF)>湖南永州(HNYZ)>福建延平(FJYP)>福建顺昌(FJSC)。
4 小结与讨论
4. 1 小 结
掌握林木的生长节律是更好地开展林木作业的前提。观光木幼苗的生长节律揭示了其在不同时期的生长特点[ 21 - 22 ]。苗高和地径是林木苗期生长的重要指标[ 23 ],不同种源观光木幼苗苗期生长发育速率具有差异性,在生长季中生长量增量变化也呈显著节律性,但生长规律大体相同。从4月整体出苗至7月期间生长较为缓慢,7月至10月下旬苗高生长速度较快,11月之后苗高生长缓慢,这与程世[ 17 ]的研究结果相似。其中赣南树木园种源(GNSMY)和南宁树木园(NNSMY)的苗高生长幅度在9~11月期间与其他种源有明显不同,依旧保持高生长速率。结合生长季气候因素与管理措施可知,虽然7~9月为夏季高温天气,降水量少,会一定程度制约观光木幼苗的生长速率,但在此期间,为了提高苗木质量保证苗木生长,加强水肥管理和病虫害防治,安全度过高温天气,极大的促进了苗木的生长。
变异系数表示性状值离散性特征,变异系数越大,性状值离散程度越大,表型多样性越丰富;变异系数小,说明该种群的性状变异幅度低。不同种源间生物量变异系数较大,表明不同种源间的遗传变异在生物量上表现得较为明显[ 24 - 26 ]。不同地理种源观光木表型性状指标、生物量均存显著差异(P<0.01)。其表型性状平均变异系数为14.11%,变异幅度为8.15%~21.10%,平均变异系数最小的是永州种源(HNYZ),最大的是南宁种源(NNSMY),表明永州种源(HNYZ)表型多样性比较丰富,可能是观光木的表型多样性中心。从生物量的变异系数上来看,观光木各种源间茎鲜重的变异系数要大于根鲜重,但整体上看,茎鲜重也高于根鲜重。说明观光木各种源间的生物量具有一定的稳定性。从种源层面来看,南宁种源(NNSMY)苗高、地径、叶长、叶宽、叶面积、茎鲜重、茎根比、叶片数量等指标在11个种源中均处于优势。总体表现出南宁种源(NNSMY)较好,其他种源生长势稍差,但是生长表现良好,均能适应栽植地的环境。
观光木子代幼苗表型性状与种源生态地理的相关性分析表明。种源和经度影响观光木苗木地上部分呈显著负相关,叶绿素和碳含量与种源、经度、海拔均呈显著正相关,但与纬度呈显著负相关。地理因子与观光木苗期各表型性状的相关性绝大部分为显著负相关,表明地理因子对1年生观光木幼苗表型性状的影响存在一定的差异性。说明观光木在低纬度、温度高、降水充沛的地方生长良好。
通过聚类分析将11个种源划分为3类,第三类南宁树木园种源为优良种源,该种源的苗高、地径、叶长、叶宽、叶面积平均值均为最大。通过聚类分析,可以发现苗期性状在各种源间的表现存在差异,以及各种源在不同性状上展现的独特优势,进一步缩小了对目标性状的选择范围,有助于对各观光木种源的筛选。采用隶属函数法对11个地理种源观光木幼苗的优良度进行综合评价,结果表明,南宁种源(NNSMY)、始兴种源(GDSX)的苗木生长较好,而延平种源(FJYP)、顺昌种源(FJSC)生长较差,其他种源介于两者之间。
4. 2 讨 论
任何一种生物其早晚生长存在密切联系,利用幼成相关评价可以估测这些贡献在后发育阶段的表现。因此,开展不同地理种源苗期生长变异研究,可以提早的筛选出优良种源。本研究表明,不同地理种源1年生观光木苗木表型性状和生物量均存在显著差异,说明观光木优良种源在苗期就有较大的选择潜力,这与前人对野鸦椿[ 27 ]、麻栎[ 28 ]、闽楠[ 29 ]等树种种源苗期的研究结果一致。本研究中11个种源分布在4省11县,但广西、湖南省仅有1个种源,与其他两省种源的差异性还有待进一步探讨。
在木本植物中,复杂的环境条件、长期的地理隔离和自然选择容易引起种内表型变异,同时表型性状的变异也受到遗传和环境因素影响[ 30 ]。观光木子代幼苗表型性状以及生物量在不同种源间的呈现,揭示了1年生观光木幼苗种间的差异在表型性状和生物量显著表现,从两者的变异系数来看,各种源间表型性状的变异系数变化幅度较小,表型性状间的变化幅度较大,生物量的变化幅度较大,即不同地理种源1年生观光木幼苗的差异主要由表型性状和生物量决定。然而随着观光木幼苗的生长及上山造林后,表型性状和生物量又会如何变化,种源间的差异性是否依然显著,种源地理因子和气候对观光木的影响程度,这都有待于后期研究。
目前,前人对不同种源苗期性状变异的研究已取得一定的成果。但在进行种源评价和筛选时,选取的指标过于单一,且多考虑地上部分生长,忽略了地下部分的重要性。本文采用隶属函数法,综合考虑表型性状和生物量主要指标的贡献,筛选出南宁树木园种源为苗期生长表现较好的种源。该种源的地上部分苗高、地径、叶长、叶宽、叶面积平均值最大,地下部分主根长、茎鲜重、根鲜重指标在均处于前列。可见,进行观光木种源苗期初选时,也要考虑地下部分的重要性。苗期在优良种源的选择中只是一个开端,不同种源植株后期生长是否与苗期生长具有一致性,还需要在造林后进行更深层次的观测研究。
参考文献
[1] 陈晓阳, "沈熙环. "林木育种学[M]. "北京: "高等教育出版社, 2005.
[2] 张兰. "不同种源棕榈种苗生物学特性变异及初步评价[D]. 贵阳: 贵州大学, 2017.
[3] 陈澜. "不同种源福建柏苗期性状差异性研究及优良种源选择[D]. 福州: 福建农林大学, 2015.
[4] 杨宗武, "郑仁华, "肖祥希, "等. "福建柏种源苗期生长和根系性状的遗传变异[J]. 南京林业大学学报(自然科学版), 2001, 3: 26 - 30.
[5] 王戈, "唐源盛, "杨汉波, "等. "桢楠优良种源/家系苗期评价和选择研究[J]. 四川林业科技, 2019, 40(3): 63 - 66.
[6] 陈莹莹. "火力楠种源区域试验与优良种源早期选择[D]. 南宁: 广西大学, 2018.
[7] 黄琛. "油松优良种源的早期选择[D]. 秦皇岛: 河北科技师范学院, 2018.
[8] 胡文杰, "王晓荣, "胡兴宜, "等. "枫香优良种源及家系早期选择[J]. 东北林业大学学报, 2017, 45(6): 5 - 11.
[9] 林正聪. "马褂木种源评价及优良种源早期选择[D]. 南宁: 广西大学, 2016.
[10] 覃敏. "米老排优良种源/家系选择与遗传变异研究[D]. 北京: 中国林业科学院, 2016.
[11] 袁显磊. "核桃楸优良种源和家系早期选择及苗期环境因子影响评价[D]. 哈尔滨: 东北林业大学, 2013.
[12] 蒋燚, "王以红, "邱凤英, "等. "大叶栎优良种源早期选择研究[J].西部林业科学, 2011, 40(1): 1 - 7.
[13] 邱凤英,章挺,何小三,等.樟树优良种源苗期选择初级[J].湖南林业科技,2021,48(5):1 - 6.
[14] 胡德活, "阮梓材, "卓铜勋, "等. "杉木优良无性系早期选择[J].广东林业科技, 1998(3): 7 - 12.
[15] 郑畹, "舒筱武, "冯弦. "云南松优良种源生长量早期选择的研究[J]. 云南林业科技, 1998, 3: 13 - 18.
[16] 池毓章. "观光木播种苗生长规律及育苗技术研究[J]. 福建林业科技, 2007(1): 122 - 125, 132.
[17] 程世, "饶玮, "张艳杰, "等. "观光木一年生播种苗生长发育规律及育苗技术研究[J]. 现代农业科技, 2008(19): 17 - 18.
[18] 吉悦娜, "颜立红, "彭春良, nbsp;等. "观光木一年生苗生长规律研究[J]. 湖南林业科技, 2008(3): 17 - 19.
[19] 丁梦军, "郝丽珍, "张凤兰, 等. "内蒙古5种蒲公英营养成分分析[J]. 饲料工业, 2015, 36(21): 20 - 24.
[20] 王付刚. "脂松种源试验和樟子松生长对比分析[J]. 绿色科技, 2017(7): 22 - 24.
[21] 肖祖飞, "艾卿, "金志农, "等. "芳樟矮林生长节律及精油动态变化研究[J]. 江西农业大学学报, 2021, 43(4): 834 - 841.
[22] 于艳, "李立峰, "费滕, "等. "人工抚育对蒙古栎幼苗成活率及生长量的影响[J]. 北方蚕业, 2021, 42(1): 36 - 38, 42.
[23] 何庆海, "方茹, "李文鑫, "等. "不同种源枫香树幼苗生长性状的地理变异[J]. 植物资源与环境学报, 2019, 28(2): 88 - 95.
[24] 李鑫, "李昆, "段安安, "等. "不同地理种源云南松苗期表型性状与生物量比较研究[J]. 西北林学院学报, 2020, 35(1): 94 - 100, 139.
[25] 明军, 顾万春. "紫丁香表型多样性研究[J]. 林业科学研究, 2006(2): 199 - 204.
[26] 龚梅香. "巧玲花天然群体遗传多样性研究[D]. 北京: 北京林业大学, 2007.
[27] 陈菊艳, "陈锐, "陈志萍, "等. "不同种源野鸦椿一年生苗生长规律研究[J]. 种子, 2021, 40(5): 126 - 131, 135.
[28] 王标, "虞木奎, "王臣, "等. "不同种源麻栎苗期生长性状差异及聚类分析[J]. 植物资源与环境学报, 2008, 17(4): 1 - 8.
[29] 邱生龙. "闽楠幼林不同地理种源生长差异及优良种源选择[J].武夷学院学报, 2017, 36(9): 39 - 43.
[30] 潘昕昊. "不同种源米槠种子及苗期差异性分析研究[D]. 南昌: 江西农业大学, 2022.