摘要:" 为明确黑龙江省黄檗天然种群的遗传分化机制,揭示不同地区黄檗天然种群的遗传关系,本研究利用 SSR 分子标记对325份来自黑龙江省的黄檗群体进行遗传多样性和遗传结构分析,结果表明,黄檗期望杂合度(He)为0.481,观测杂合度(Ho)为 0.422,信息指数(I)为0.965,说明黄檗遗传多样性较高,遗传分化系数为 Fst=0.067,说明遗传分化水平也处于较高状态,AMOVA分析显示,黄檗遗传变异中4%发生在种群间,83%发生在个体间,说明种间变异小;对黄檗14个种群遗传结构分析,遗传分化系数p=0.031<0.05,说明黄檗分布在不同种群之间存在显著的遗传分化。
关键词:" 黄檗; 遗传多样性; SSR分析
中图分类号:" "Q 792. 31" " " " " " " "文献标识码:" "A" " " " " " " " 文章编号:1001 - 9499(2025)02 - 0014 - 05
SSR Analysis of Genetic Diversity of Natural Phellodendron
amurense Populations in Heilongjiang Province
YUAN Xianlei WANG Yutong ZHOU Zhijun LIU Zhongling ZHANG Jianying**
(Heilongjiang Academy of Forestry," Heilongjiang Harbin 150081)
Abstract In order to clarify the genetic differentiation mechanism of natural Phellodendron amurense populations and to reveal the genetic relationship of natural Phellodendron amurense populations in different regions, this study analyzed the genetic diversity and genetic structure of 325 Phellodendron amurense populations from Heilongjiang Province using SSR molecular markers. The results showed that the expected heterozygosity (He) of Phellodendron amurense was 0.481, the observed heterozygosity (Ho) was 0.422, and the information index (I) was 0.965, indicating that the genetic diversity of Phellodendron amurense was high. 0.965, indicating that Phellodendron amurense has high genetic diversity, and the coefficient of genetic differentiation is Fst=0.067, indicating that the level of genetic differentiation is also in a high state. AMOVA analysis showed that 4% of Phellodendron amurense genetic variations occurred between populations, and 83% of the genetic variation occurred between individuals, indicating that interspecific variations were small; the analysis of 14 Phellodendron amurense populations of genetic structure, and the coefficient of genetic differentiation, P=0.031<0.05, indicated that Phellodendron amurense was distributed in different populations. This indicates that Phellodendron amurense is distributed among different populations with significant genetic differentiation.
Key words Phellodendron amurense; genetic diversity; SSR analysis
黄檗(黄柏,黄菠萝Phellodendron amurense)作为东北三大硬阔树种之一,是重要的用材和药用树种。在用材上主要为我国东北阔叶红松林重要的伴生乔木;在药用上是我国传统中药黄柏药源植物[ 1 ]。由于人类过度开发用于中药和木材,野生黄檗种群急剧下降,遭受栖息地碎片化和丧失。因此,它被列为《国家重点保护野生植物名录》(2021)国家二级保护植物[ 2 - 3 ],由于过度的开发利用,黄檗这一野生植物已处于濒危状态,急需保护和修复。濒危物种的长期生存几乎完全取决于它在适应持续的环境变化时保持足够的遗传变异的能力[ 4 ]。利用分子标记估计遗传多样性和种群结构已成为濒危物种保护的一种常见且最有效的方法[ 5 ]。
已有研究通过利用 ISSR 标记、AFLP 标记对黄檗的遗传多样性及群体结构进行了分析[ 1 , 6 ]。有关遗传多样性方面的工作较少,以至于无法提出有针对性的保护策略。SSRs(微卫星标记)具有共显性、稳定性好、多态性高等优点[ 7 - 8 ],被广泛用于遗传多样性评价和群体遗传结构的研究。例如,AFLP标记已经被用来测量山毛松的遗传多样性[ 9 ]。简单序列重复序列(SSR)具有高度的多态、共显性、高重复性和广泛的基因组覆盖率等特点,已被证明是野生物种群体遗传学研究的高效分子工具[ 10 - 11 ]。
本研究利用 SSR 分子标记对325份来自黑龙江省地区的黄檗群体进行遗传多样性和遗传结构分析,揭示不同地区黄檗天然种群的遗传关系,探讨黄檗天然种群的遗传分化机制,旨在为黑龙江省黄檗种质资源的有效保护和高效利用提供理论依据。
1 材料与方法
1. 1 试验材料
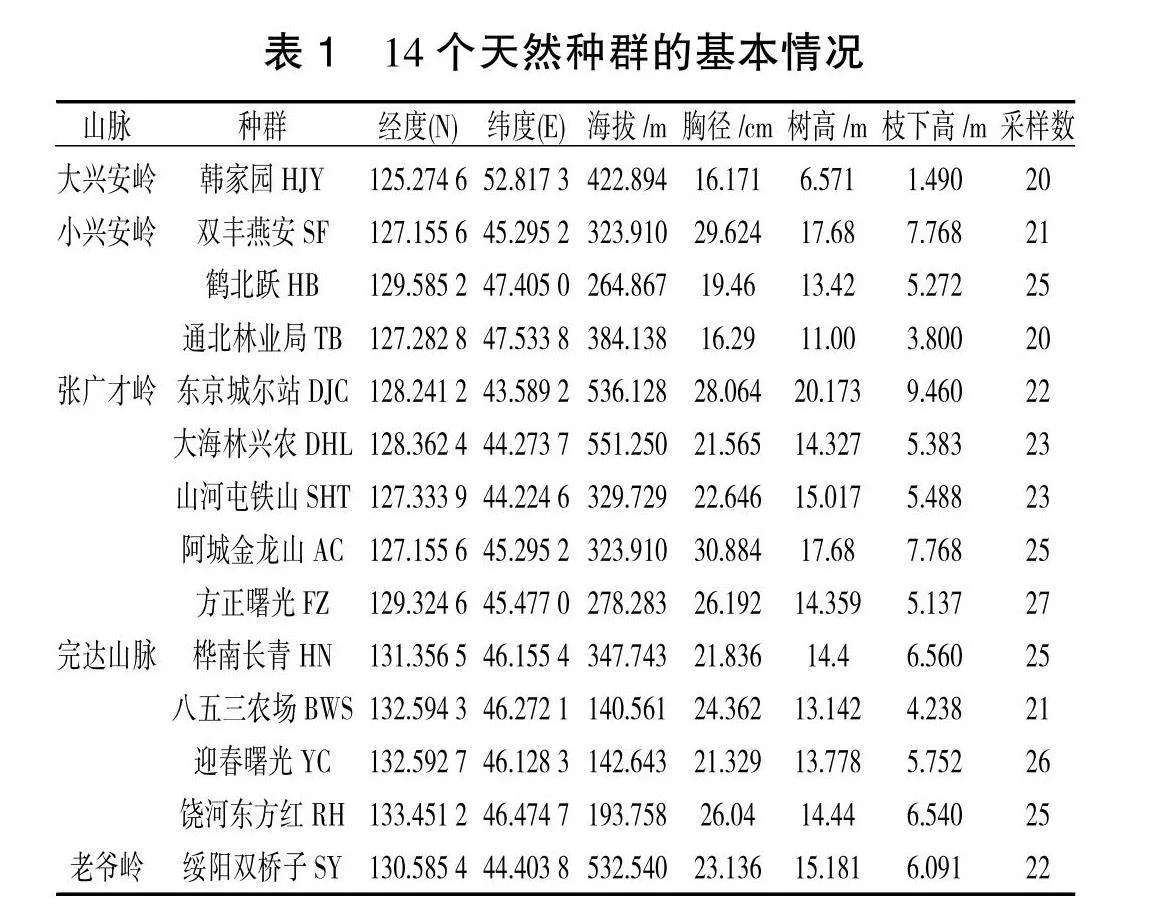
根据黑龙江省山脉地理位置及森林资源调查确定了 14个有代表性的天然种群(表1)。林分要求立地较好的近熟林分,采样时间是2023 年7月。为使样本具有代表性,各样本间相互距离要在 50 m以上,采集完整幼嫩叶片并使用硅胶干燥,放入冰盒后迅速转运实验室,储存到-80 ℃冰箱,待用。
1. 2 DNA提取与SSR扩增
利用DNA试剂盒(天根)提取黄檗叶片总DNA,从黄檗的参考文献中筛选26对 SSR引物[ 6 ]。本试验中所采用的 PCR 反应体系:95 ℃预变性5 min;95 ℃变性30 s,62~52 ℃梯度退火30 s,72℃延伸30 s,运行10个循环;95 ℃变性30 s,52 ℃退火30 s,72 ℃延伸30 s,运行25个循环;72 ℃延伸20 min,最后4 ℃保存。反应在PCR(Veriti 384well,Applied Biosystem)仪进行。PCR反应结束后,扩增产物经荧光毛细管电泳(DYY-6C,北京六一)在 ABI 3730xl DNA分析仪中检测。
1. 3 数据的统计分析
利用GenAlEx version 6.501软件,计算SSR位点和群体的各项遗传多样性指标,包括观测等位基因(Na)、有效等位基因(Ne)、香农指数(I)、多态性信息指数(PIC)、观测杂合度(Ho)、期望杂合度(He)和近交系数(Fis)。利用powermarker软件计算各群体间的遗传距离。利用UPGMA方法进行聚类分析,并绘制聚类图。利用STRUCTURE 2.3.4对325个样本进行群体结构分析。根据群体遗传结构分析结果,在GenAlEx version 6.501 软件中计算各群体间和群体内的变异、分化并进行显著性检验;计算遗传分化系数(Fst)和基因流(Nm)。
2 结果与分析
2. 1 黄檗种群遗传多样性分析
2. 1. 1 引物多态性分析
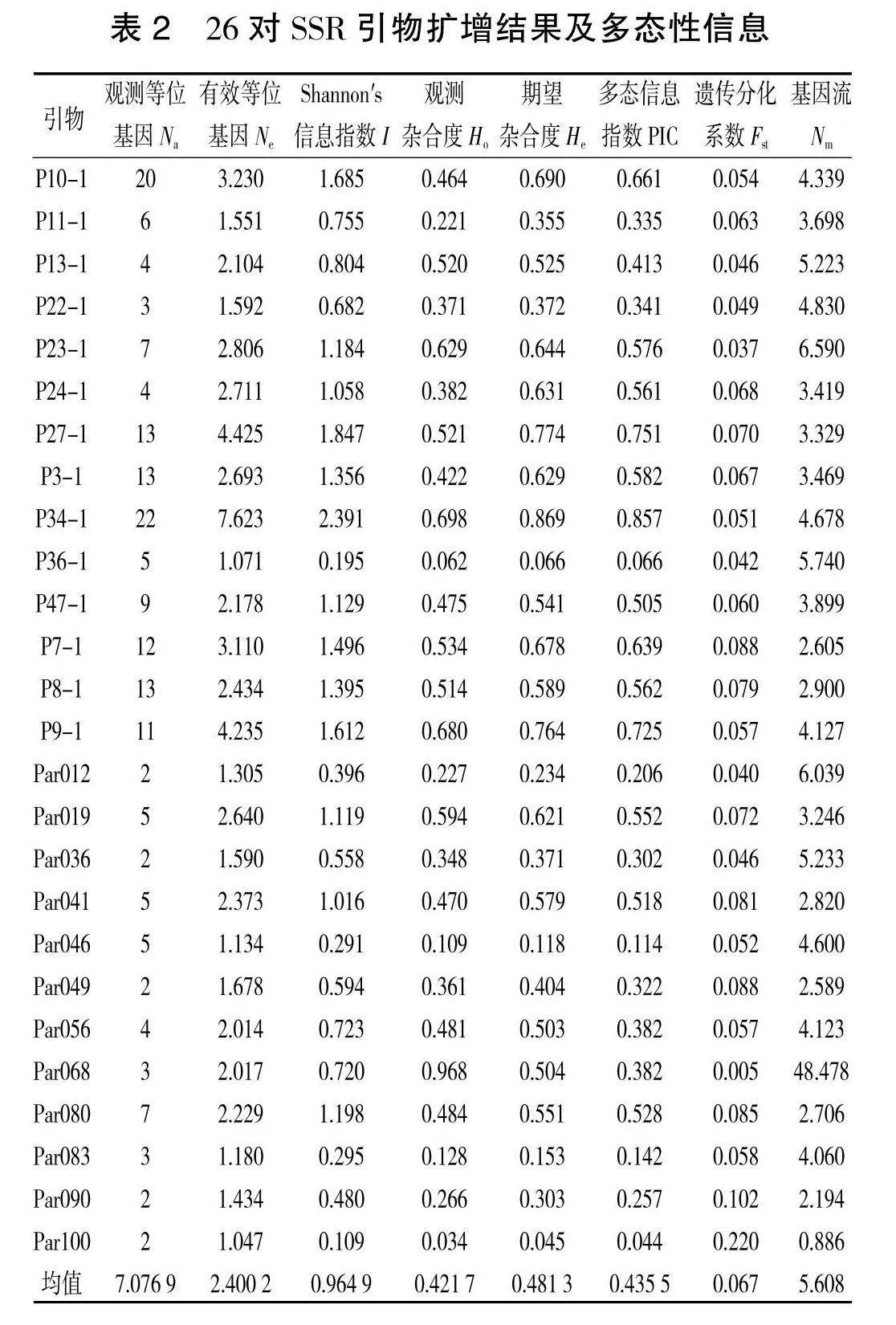
利用 26 对 SSR 引物对黑龙江省天然林下黄檗14个种群325份样本进行遗传多样性评价,SSR引物扩增结果及多态性分析显示见表2。26对SSR引物共检测出184个等位基因(Na),平均每个位点等位基因数为7.0769。在扩增出的 184个等位基因中有62.404个有效等位基因(Ne),平均每个位点有效等位基因数为2.4002。这表明黄檗不同种群下特异带和种群间共有带的某些位点上差异明显。从整体来看,供试种质的观测杂合度(Ho)为 0.034(Par100)~0.968(Par068),平均值为0.421 7;期望杂合度(He)为 0.045(Par100)~0.869(P34-1),平均值0.481 3,两者均值表现为 Holt;He,说明供试种质之间存在一定程度的近交现象,导致纯合子过量。Shannon′s信息指数(I)为0.109(Par100)~2.391(P34" -1),平均值0.9649。引物多态性信息含量(PIC)为0.044(Par100)~0.857(P34-1),平均值0.4355,其中 PICgt;0.5 的高多态性位点有13个,0.25lt;PIClt;0.5的中度多态性位点有7个;PIClt;0.25的低度多态性位点有3个。这表明这26对SSR引物在黄檗上有较高水平的扩增位点多态性不同SSR位点的多态性有明显差异。遗传分化系数(Fst)范围为 0.220~0.005。基因流 (Nm)范围为48.478~0.886。
2. 1. 2 种群的分子方差分析
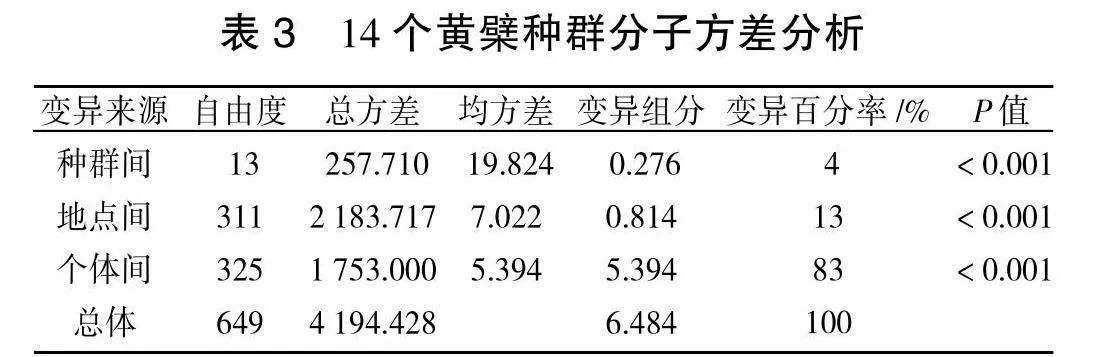
为确定14个天然种群的变异特征,遗传分子变异分析显示(表3),黄檗自然种群间遗传分化系数Fst为0.043,总遗传变异中有83%变异存在于个体内,种群间遗传变异的发生率仅为4%,且种群内和种群间差异均极显著(Plt;0.001)。分子方差分析是一种通过进化距离来度量并计算单倍型(或基因型)间遗传变异的方法。分子方差分析表明,4%的遗传变异存在于居群,有96%的遗传变异存在于个体,个体的变异是总变异的主要来源。
2. 1. 3 种群遗传多样性分析
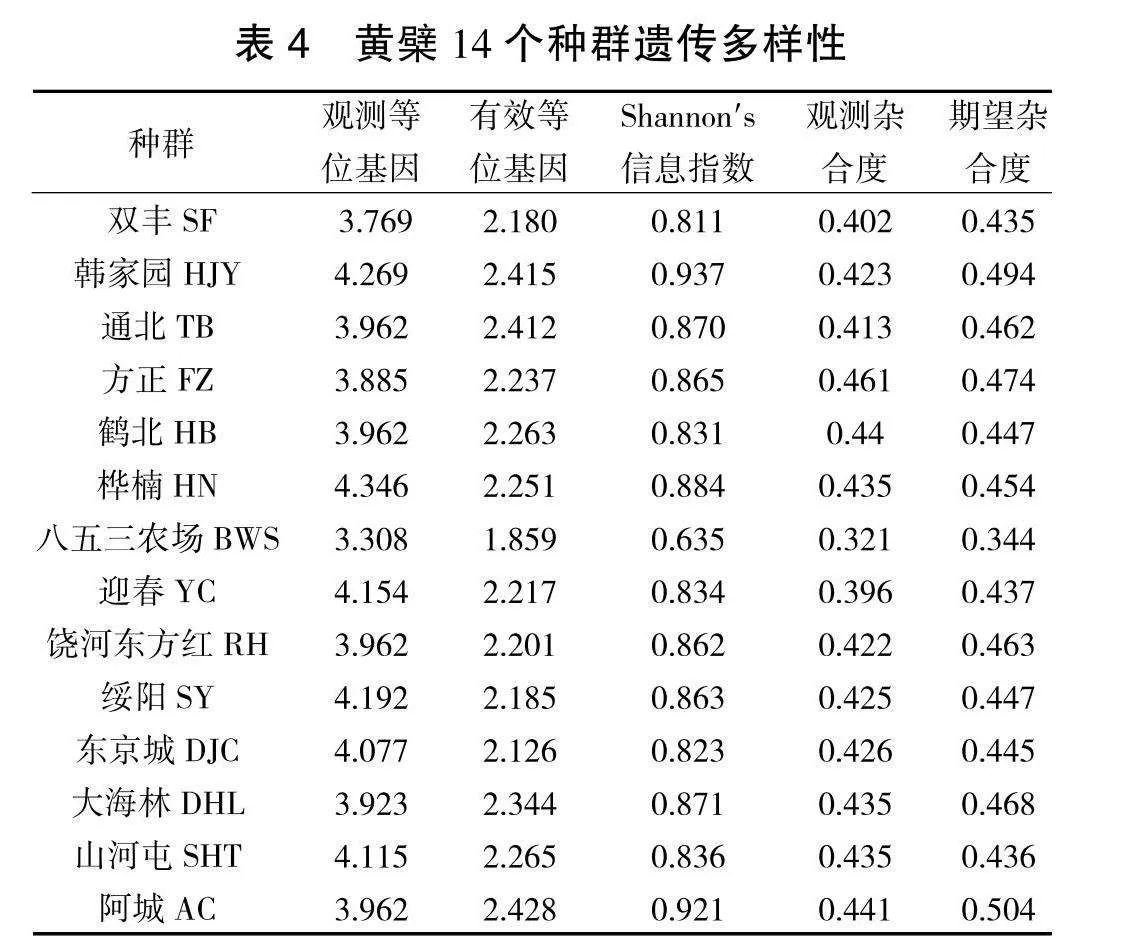
遗传多样性结果(表4)表明,等位基因数(Na)范围为4.346(HN)~3.308(BWS)。有效等位基因数(Ne)范围为2.428(AC)~1.859(BWS)。信息指数(I)范围为0.937(HJY)~0.635(BWS)。观测杂合度(Ho)范围为0.447(HB)~0.321(BWS)。期望杂合度(He)范围为0.504(AC)~0.344(BWS)。综合来看,小兴安岭种群的遗传多样性水平最高,完达山脉种群的遗传多样性水平最低。
2. 2 黄檗遗传结构分析
2. 2. 1 种群间主成分分析
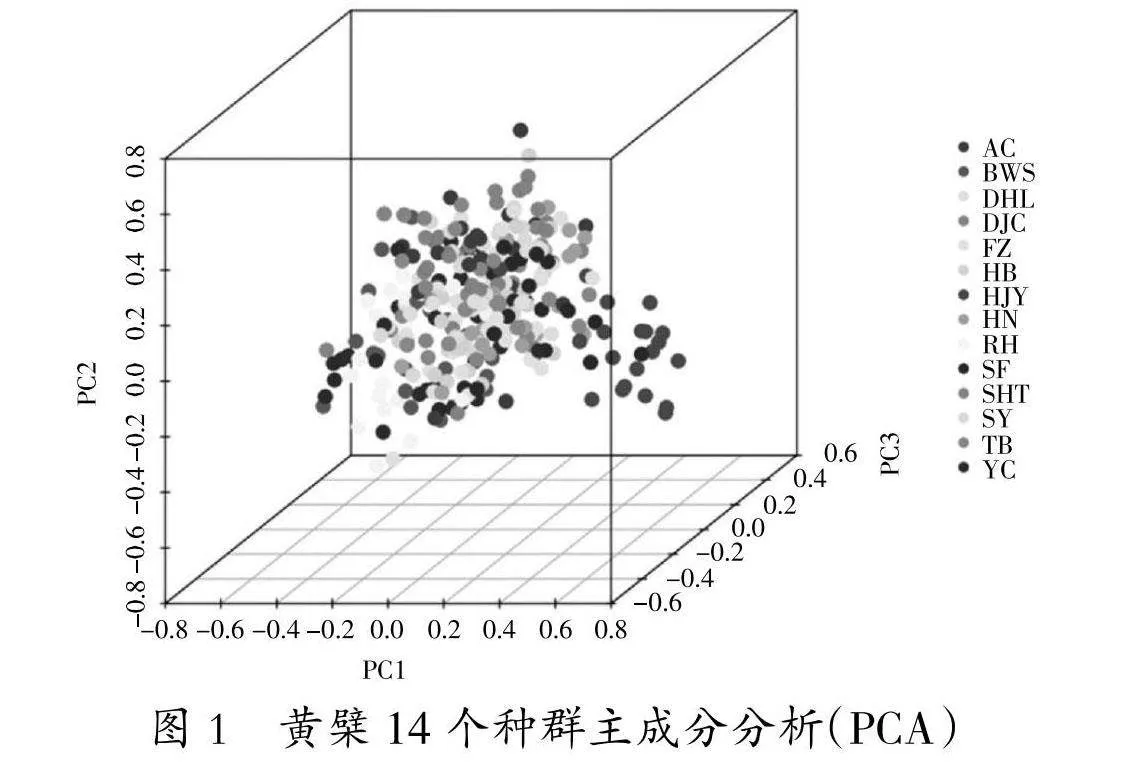
利用 GenAlEx6.5进行主成分分析(图1),他们在图中的距离代表亲缘关系的距离,三个坐标的贡献率分别为5.95%、4.55%和4.27%。14个种群分成2个类群:第1类群包括HJY、YC、RH、BWS共4个种群;第2类群包括 FZ、SY、HN、HB、SF、TB、DHL、AC、SHT、DJC 共 10个种群。在中心位置有高度重合,各种群间差异性小,其中HJY差异最大,亲缘关系最远。
2. 2. 2 种群聚类分析
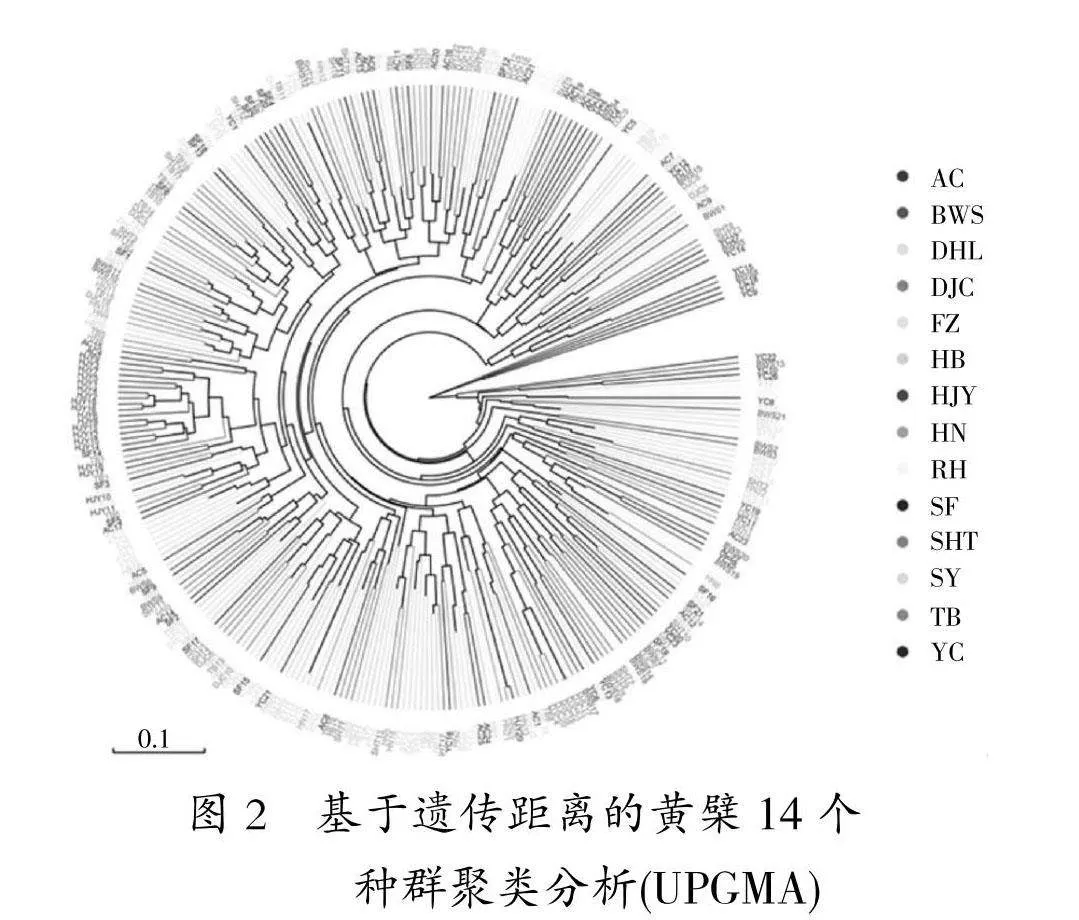
14个群体间遗传距离最大为0.179" 549 (YC/HJY),最小为0.044 252 (SHT/AC),对黄檗14个种群进行基于遗传距离的UPGMA 聚类分析(图2),与主成分分析结果一致。
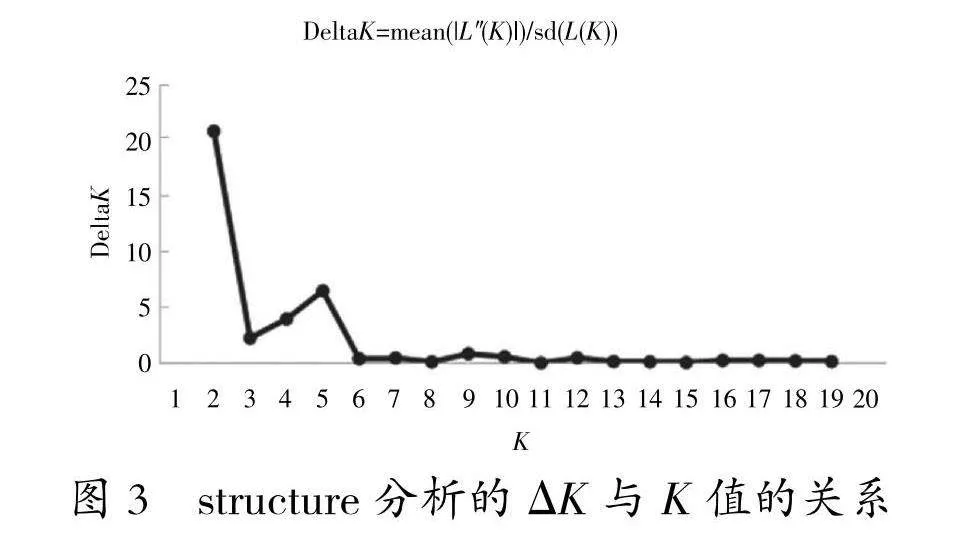
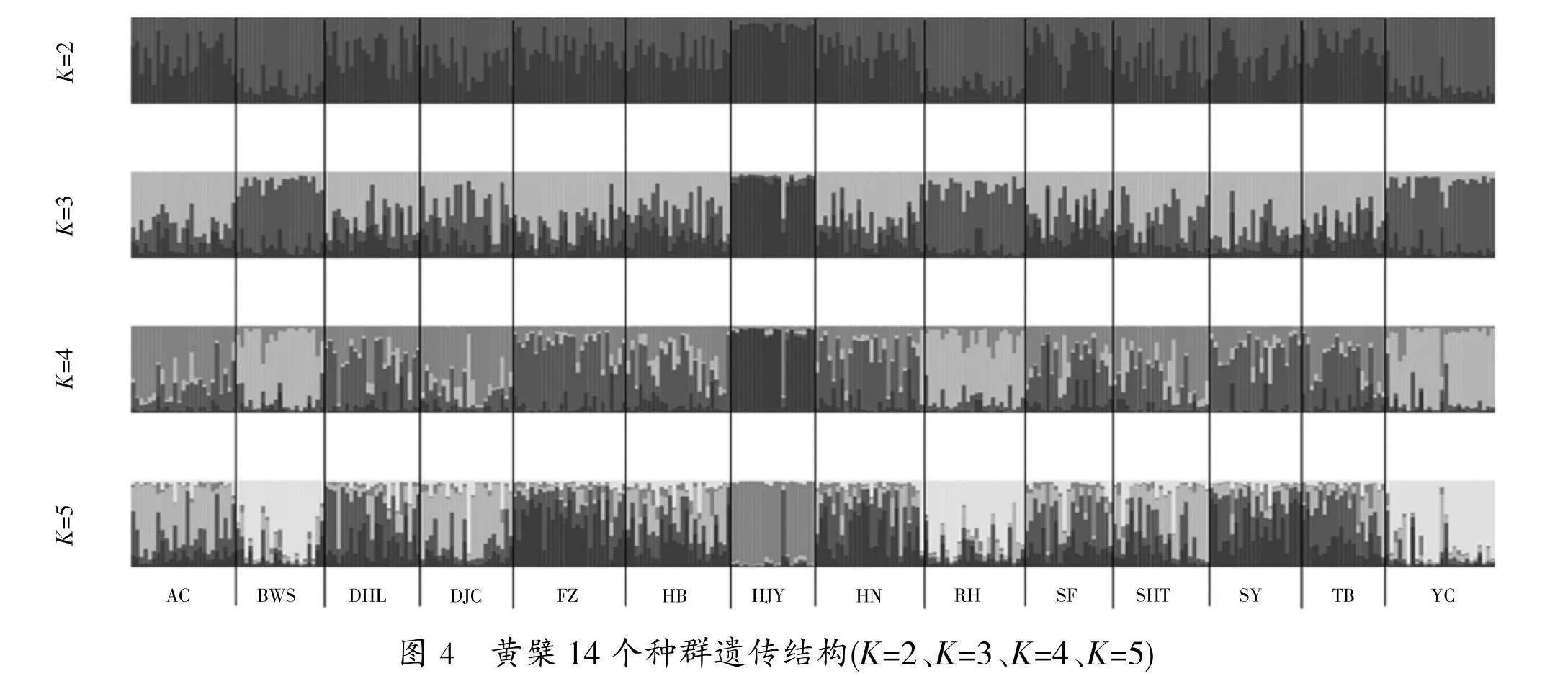
对黄檗14个种群325份种质资源进行了遗传结构分析(图3,图4)发现,当K=2时,ΔK达到最大值,说明遗传变异来源于2个基因池,可以将325份材料分为2个亚群,亚群Ⅰ、亚群Ⅱ,分别包含了158份、167份种质。黄檗14个种群遗传分化系数P=0.031<0.05,表明黄檗分布在不同种群之间存在显著的遗传分化。
3 讨 论
20世纪60年代开始东北林区长期大规模砍伐黄檗加工木材加上盗剥树皮,野生资源遭到严重破坏,导致黄檗的成林面积逐渐减少,至今种群规模未得到较好的恢复。目前野外已经很少有成片分布的黄檗天然林[ 13 ]。分子方差分析是一种通过进化距离来度量并计算单倍型(或基因型)间遗传变异的方法。本研究种群AMOVA分析显示,黄檗遗传变异中4%的遗传变异存在于居群,有96%的遗传变异存在于个体,说明个体的变异是总变异的主要来源。黄檗分布区内随着山脉的走向的气候条件差异显著,尤其大兴安岭地区遗传距离最远,黄檗天然林长势最弱。黄檗花为单性且雌雄异株,天然黄檗的繁殖主要靠种子繁殖,并依靠鸟类的采食把种子传播到较远的地方,通过异花授粉促进了个体间的基因交流、增加了有效种群的大小、减少了基因漂变对遗传结构的改变[ 14 ]。在实地调查过程中发现,种群个体间差异很大,发现了个别优良性状或者特殊性状的黄檗植株。这样的黄檗个体间丰富性差异可以为遗传改良提供了有效资源。但是,目前黄檗天然种群的更新能力极差,现存的黄檗种质资源流失严重,相互隔离的小种群使黄檗的遗传变异性更加减少,使这些种群也存在较高的灭绝风险。
利用SSR 荧光标记具有构建林木种质分子身份证和鉴别雌雄株等优势[ 15 - 16 ],本研究利用SSR荧光标记后,从整体上看黄檗期望杂合度(He)为0.481,观测杂合度(Ho)为 0.422,信息指数(I)为0.965,说明黄檗遗传多样性较高,遗传分化系数为 Fst=0.067,遗传分化水平也处于较高水平。有限的基因流是居群形成空间遗传结构的主要原因。遗传变异对任何物种保持其进化潜力以应对不断变化的环境的能力起着至关重要的作用。因此,保持遗传变异是大多数濒危物种保护计划的主要目标[ 17 - 20 ]。但基于本研究的结果,黄檗保持了较高的遗传多样性,所以该物种濒临灭绝的主要原因可能是对野生种群的过度开发,而不是缺乏整体遗传多样性。因此,必须制定政策,禁止在野外非法砍伐的同时提高人工栽培力度,以提高生态效益和经济效益。
参考文献
[1] 李绍臣, 李凤明, 张立民, 等. 吉林省天然黄檗种群遗传多样性ISSR分析[J]. 生态学报, 2016, 36(13): 4006 - 4012.
[2] 傅立国. 中国珍稀濒危保护植物名录[M]. 北京: 科学出版社,1992.
[3] 方文培, 张泽荣. 中国植物志: 第 52 卷: 第 2 分册[M]. 北京:科学出版社, 1983: 94.
[4] COTE C T. Genetic variation in rare and common plants[J]. Annu. Rev. Ecol. Syst. 2003,34(5): 213 - 237.
[5] GROOM M J, MEFFE G K, CARROLL C R. Principles of Conservation Biology[M]. USA: Sinauer Associates, Inc., 2006.
[6] 陈常美. 东北林区主要珍贵用材树种种质遗传多样性评价与保护策略[D]." 哈尔滨: 东北林业大学, 2013.
[7] 程玮哲, 樊军锋, 周永学. 基于荧光 SSR 标记的10个白杨 派种质资源遗传多样性分析[J].西北林学院学报, 2021, 36(3):88 - 93.
[8] 姚建忠. 中国北方青杨派杨树遗传多样性研究[J]. 西北林学院学报, 2024, 39(2): 79 - 84.
[9] 闫志峰, 张本刚, 张昭, 等. 珍稀濒危药用植物黄檗野生种群遗传多样性的 AFLP 分析[J]. 生物多样性, 2006, 14( 6) : 488 - 497.
[10] LI X L, LI S C, CHU H J, et al. Genetic diversity and population structure of the endangered alpine quillwort Isoetes hypsophila (Isoetaceae) revealed by SSR analysis[J]. Biochem. Syst. Ecol. 2013," "47(2): 11 - 20.
[11] MENG F J, LIU L, PENG M, et al. Genetic diversity and" population structure analysis in wild strawberry (Fragaria nubicola L.) from Motuo in Tibet Plateau based on simple sequence repeats (SSRs) [J]. Biochem. Syst. Ecol. 2015, 63(1): 113 - 118.
[12] HAMRICKJL. Isozymes and the analysis of genetic structure in plant populations[J]. Plant Biology,1989(2): 87 - 105.
[13] 张志鹏, 张阳, 张昭. 我国黄檗野生种群生存现状及化学表征研究[J]. 植物科学学报, 2016, 34(3): 381 - 390.
[14] 闫平玉, 张磊, 王佳兴, 等. 红松天然种群遗传多样性分析及核心种质构建[J]. 南京林业大学学报(自然科学版), 2024, 1 - 17.
[15] 秦宇, 郭荣琨, 农惠兰. 利用SSR荧光标记构建山楂种质分子身份证[J]. 园艺学报, 2024(3): 1 - 14.
[16] 齐凤慧, 孙宏冉, 詹亚光. EST-SSR标记在水曲柳雌雄鉴定中的应用[J]. 西北植物学报, 2015, 35(3): 472 - 479.
[17] AVISE J C, HAMRICK J L. Conservation Genetics: Case Histo- ries from Nature[M]. New York, USA: Chapman and Hall,1996.
[18] HAMRICK J L, GODT M J W. Effects of life history traits on" " genetic diversity in plant species[J]. Philos. Trans. R. Soc. Lond. B 1996, 351(4): 1291 - 1298.
[19] 刘宝富. 13份油茶种质资源的表型遗传多样性分析[J]. 安徽农业科学, 2022, 50(23): 93 - 95.
[20] 游书梅, 曹应江, 郑家奎, 等. 73份亚洲水稻恢复系农艺性状的主成分与聚类分析[J]. 植物遗传资源学报, 2015, 16(2): 250 - 256.