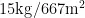摘 要:【目的】分析无核白和芽变长粒无核白(长粒无核白)葡萄之间的植物学性状、果实品质、葡萄干多酚和抗氧化活性的差异,为葡萄育种提供新的种质资源。
【方法】以无核白葡萄及芽变长粒无核白为试材,调查植物学性状、光合响应曲线、果实品质和葡萄干品质差异,分析葡萄干酚类物质和抗氧化活性之间的关系。
【结果】长粒无核白葡萄主要变异特征表现在嫩叶、嫩梢、叶柄颜色和果粒上。与普通无核白相比,长粒无核白葡萄幼叶正面颜色绿色带红条带,新梢腹侧颜色绿带红条带、节间背侧颜色绿带红条带、节间腹侧绿带红条带。无核白萌芽率高于长粒无核白,其萌芽率分别为65.00%(2022年无核白)、63.82%(2023年无核白)、50.48%(2022年长粒无核白)、53.45%(2023年长粒无核白)。在整个果实发育期间,长粒无核白的果形指数始终显著高于无核白,花后16 d无核白葡萄的果形指数最大是1.80,而长粒无核白果实的果形指数在花后28 d达到最大,为2.12。长粒无核白果粒质量、果实还原糖含量、糖酸比显著高于无核白。无核白和长粒无核白果穗质量、果穗长度、果穗宽度、果实可溶性固形物、可滴定酸、VC和果皮色差差异不明显。无核白的出干率、平均质量均显著高于长粒无核白,而长粒无核白的果形指数(3.10)显著高于无核白葡萄干(2.13)。长粒无核白葡萄干的总黄烷醇、总类黄酮、总单宁、总酚显著高于无核白葡萄干,长粒无核白葡萄干的含量分别是1 808.59、2 286.43、51.11和498.53 mg/kg。长粒无核白葡萄干的DPPH自由基清除能力、对Fe2+的还原能力显著高于无核白葡萄,长粒无核白葡萄干的ABTS自由基清除能力低于无核白葡萄干,但无显著差异。葡萄干总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力、FRAP(对Fe2+的还原能力)彼此之间的相关系数大于0.926,为显著或极显著差异,而ABTS和总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力和FRAP(对Fe2+的还原能力)呈负相关,但无显著差异。
【结论】生产上芽变长粒无核白葡萄可以作为优良品种开发利用,亦可为育种提供新的种质资源。
关键词:无核白;长粒无核白;芽变;果实品质;葡萄干
中图分类号:S663.1"" 文献标志码:A"" 文章编号:1001-4330(2024)12-2954-12
0 引 言
【研究意义】无核白(V.vinifera cv.Thompson Seedless)属欧亚东方品种群,中晚熟品种,是一个以制干为主的多用途优良葡萄品种[1]。新疆吐鲁番市葡萄栽培面积近3.36×104 hm2,其中90%以上为无核白葡萄,无核白葡萄干年产量超过14×104 t,无核白葡萄产业是该区农业主导产业[2-3]。芽变是体细胞突变的一种,即体细胞在芽的分生组织细胞中发生突变并经过萌发长成枝条,其性状表现不同于原类型的现象[4]。葡萄是最易产生芽变的果树,也是通过芽变选育品种最多的一类果树。【前人研究进展】20世纪50年代末,在新疆鄯善县汗墩发现了无核白芽变系长无核白葡萄,1964年从芽变株上剪条育苗,1983年在新疆葡萄瓜果开发研究中心(现新疆维吾尔自治区葡萄瓜果研究所)育苗定植(4亩)800余株,1984年经石河子农学院园林系鉴定根尖染色体为2n=38[5]。除长粒无核白外,无核白优良芽变系还有长穗无核白[3]、大粒无核白[6]。长粒无核白葡萄生长结果习性与无核白相似,其葡萄干果形状为柱形,碧绿-绿黄色,外形美观,葡萄干单粒质量较无核白大,口感、饱满度均与无核白相似,综合品质优于无核白。闫鹏等[7] 不仅对其植物学形态特征观察,还利用RAPD分析了其基因突变及遗传多样性;张宗勤等[8]分析比较了无核白葡萄营养系果实的酚类物质差异,长粒无核白果实的白藜芦醇含量较高,但酚类物质含量较低;别尔麦提布·依明等[9]从生物学性状、光合特性、结实特性分布规律等方面进行了比较分析,选区1、选区4、选区5、选区6芽变区可能为优异变异单株,并任务长粒无核白葡萄丰产性较强,可扩大栽培面积;薛锋等[10]分析了吐鲁番市5个无核品种的栽培特性、果实品质和制干特性差异,葡萄综合品质排序为紫甜无核gt;无核白鸡心gt;长粒无核白gt;波尔莱特gt;无核白,葡萄干综合品质排序为无核白鸡心gt;紫甜无核gt;长粒无核白gt;波尔莱特gt;无核白,长粒无核白葡萄品质一般,但丰产稳产性较好;户金鸽等[11]对无核白葡萄及变营养系进行了果实品质及制干特性初步分析,长粒无核白营养系品质优良。【本研究切入点】经过长期自然环境选择,长粒无核白虽然是无核白的芽变优株,但也有部分性状发生改变。需观察其植物学性状和果实性状,分析芽变长粒无核白和普通无核白品种之间植物学性状、果实品质、葡萄干酚类物质及抗氧化活性的差异。【拟解决的关键问题】研究无核白和芽变长粒无核白葡萄品质的差异性,为长粒无核白葡萄作为优良品种积极开发利用提供参考。
1.1 材 料
以新疆维吾尔自治区葡萄瓜果研究所资源圃内保存的无核白及其芽变长粒无核白葡萄为试材。株行距0.5 m×4.0 m,短梢或极短梢修剪,棚架栽培。该资源圃自1979年开始收集种质资源,80年代初步建成,2003年第1次更新保存,2018年,第2次更新保存。
1.2 方 法
1.2.1 试验设计
2022年于葡萄出土后关键时期,观察幼叶、嫩梢、叶片等植物学性状。花后10 d开始调查果形指数,直至果实成熟。果实发育期间,果实不进行植物生长调节剂处理。果实始熟期,测定光合响应曲线。待果实成熟时,测定果实品质,将剩余的葡萄用于制干试验,测定葡萄干酚类物质和抗氧化活性等生理生化指标。
1.2.2 测定指标
1.2.2.1 植物学性状
葡萄出土萌芽后,根据中华人民共和国农业行业标准(NY/T 2653-2014)《植物新品种特异性、一致性和稳定性测试指南葡萄》[12],调查无核白及芽变长粒无核白表型性状差异(萌芽率、幼叶颜色、成龄叶片性状、嫩梢、果实成熟期果实品质)。
1.2.2.2 光合响应曲线
果实始熟期,选择清朗无风的天气,分别选取长势一致的葡萄各3株,选取完全展开、叶龄一致的功能叶,提前将叶片表面擦拭干净,确保叶片在树上的原位置,采用CIRAS-3便携式光合作用测定仪(美国PP systems, CIRAS-3,美国)的开放气路(空气流速500 μmol/s),设定叶室CO2浓度为400 μmol/mol、设置光合有效辐射(PAR)(红蓝光源)梯度依次为2 500、2 200、1 900、1 600、1 300、1 000、750、500、350、200、150、100、50和0 μmol/(m2·s),测定各处理的净光合速率。将光强在0~250 μmol/(m2·s)、净光合速率(Pn)的拟合值进行直线回归,得到的直线斜率为表观初始量子效率。光相应曲线的数据采用叶子飘机理模型进行拟合,得到光合特征参数。
1.2.2.3 果形指数
葡萄花序开花坐果后1周左右开始用游标卡尺测定果粒的纵径、横径(mm),计算果形指数(果粒纵径/果粒横径),每7 d测定1次。
1.2.2.4 果实品质
果实成熟时测定果粒质量、可溶性固形物、可滴定酸、还原糖和VC。果粒质量(g)用电子天平称量,可溶性固形物(°Brix)用手持测糖仪测定,可滴定酸(%)用酸碱滴定法测定[13],VC(mg/100 g)用钼蓝比色法测定[13],还原糖(mg/L)用蒽酮比色法测定[14]。各指标重复3次。
1.2.2.5 果实色差
果实成熟时,随机选取15粒有代表性的果实用色差仪测定果皮L、a、b值,计算a/b、C、h°和ΔE。根据Hunter Lab表色系统,其中L值表示系统的亮度,L越大,样品表面越亮。a表示系统的红绿值,-a为绿,a越小样品越绿,+a为红,a越大,样品越红;b表示系统的黄蓝值,-b为蓝,值越小样品越蓝,+b为黄,值越大,样品越黄;C表示样品的彩度,值越大,表示所测的颜色越纯。C=(a2+b2)1/2;h°=arctangent(b/a),h°表示样品的色调角,h°变化幅度一般在0°~180°,h°=0°为紫红,h°=90°为黄色,h°=180°为绿色。ΔE=(L2+a2+b2)1/2,ΔE表示颜色差别大小[15]。
1.2.2.6 葡萄干的晾制及品质
葡萄干晾制:葡萄成熟时,分别选取1 500 g葡萄用促干剂(新疆惠普园艺新科技公司生产,新疆农业科学院园艺作物所监制,每包净重350.00 g,兑水15.00 kg)药液浸渍1 min,取出沥干水分,在新疆维吾尔自治区葡萄瓜果研究所晾房内制干。晾干过程中经常翻动葡萄干,防止堆积造成葡萄干腐烂变质。待葡萄干含水量降至15.00%时收集,测定葡萄干品质。测定前,挑选无病虫害,无机械损伤,色泽一致的葡萄干,用自来水将表面的杂质冲洗干净后于通风处自然晾干。
葡萄干质量、纵径、横径的测定同上。
可溶性固形物、可滴定酸、还原糖和VC含量:取20.00 g葡萄干,加蒸馏水至100.00 mL,浸泡24 h后用小型榨汁机榨至匀浆,取1.00 mL匀浆加入4.00 mL蒸馏水混匀,用手持测糖仪测定可溶性固形物(PAL-1, Atago, Tokyo, Japan);可滴定酸用酸碱滴定法测定[13];VC含量用钼蓝比色法测定 [13];还原糖用蒽酮比色法测定[14],各数值再乘以稀释倍数。
酚类和抗氧化活性的提取。取20.00 g葡萄干,加入60% HCl-CH3OH(含0.1% HCl)至 100.00 mL,待葡萄干完全浸泡透无硬心后,用小型榨汁机榨至匀浆。置于超声波清洗器(KQ3200DVDV,江苏昆山)中提取,超声提取条件:25℃、40 Hz、30 min,提取结束后8 000 r / min、4℃ 离心15 min,收集上清提取液,用于测定酚类物质和抗氧化活性。
酚类物质:单宁用Folin-Denis法测定[16],总酚用Folin-Ciocalteus法测定[17],黄烷醇用β- DMACA-HCl法测定[18],类黄酮用AlCl3比色法测定[19]。
抗氧化活性:DPPH和ABTS的测定参照张妍等[20]方法。FRAP的测定通过FRAP法[21]测定葡萄干对铁离子的还原能力。
1.3 数据处理
数据采用Excel 2010计算平均值±标准误(SD),显著性分析采用OriginLab OriginPro 2019 b (Micro Lab, Northampton, MA, USA)统计软件,在显著水平为5%条件下比较。
2 结果与分析
2.1 无核白和长粒无核白葡萄性状差异
研究表明,与无核白相比,长粒无核白葡萄发生变异的地方主要表现在萌芽始期、萌芽率、幼叶(正面颜色)、嫩梢(梢尖匍匐绒毛花青苷显色强度)、新梢(节间腹侧颜色、节间背侧颜色、节腹侧颜色、节背侧颜色)。长粒无核白葡萄萌芽始期早于无核白3~4 d(2022年3 d,2023年4 d),萌芽率(65.00%、63.82%)高于长粒无核白(50.48%、53.45%),幼叶正面颜色绿色带红条带,新梢腹侧颜色绿带红条带、节间背侧颜色绿带红条带、节间腹侧绿带红条带。表1
2.2 无核白和长粒无核白葡萄叶片净光合速率的光响应及参数差异
研究表明,叶子飘机理模型很好地拟合了净光合速率(Pn)对光合有效辐射(PAR)的响应,(R2gt;0.9),无核白高达0.994 9,长粒无核白是0.923 04。无核白和长粒无核白光响应曲线差异较大。表2,图2
长粒无核白葡萄的光饱和点(2 029.79 μmol/(m2·s))高于无核白(1 085.58 μmol/(m2·s)),较无核白增加了86.98%,无核白葡萄的光补偿点是10.53 μmol/(m2·s),高于长粒无核白的6.78 μmol/(m2·s),无核白葡萄的最大净光合速率(36.96 μmol/(m2·s))明显高于长粒无核(6.14 μmol/(m2·s)),是长粒无核白的6.02倍。无核白葡萄的表观量子效率(0.11%)明显高于长粒无核白葡萄(0.02%),无核白的暗呼吸速率是1.72 μmol/(m2·s),明显高于长粒无核白0.59 μmol/(m2·s)。表2
2.3 无核白和长粒无核白葡萄果实性状差异
2.3.1 无核白和长粒无核白葡萄果形指数的差异
研究表明,果实发育期间,长粒无核白和无核白葡萄的果粒纵径横径随果实的生长发育而缓慢增加,果形指数逐渐下降并趋于稳定。果实发育前期(花后40 d前),长粒无核白葡萄的果粒纵径始终显著高于无核白,花后40 d,长粒无核白的果粒纵径略大于无核白,但无显著差异,果实发育后期,长粒无核白的果粒纵径又开始显著高于无核白,果实成熟时,两者的果粒纵径再次达到无显著差异。长粒无核白的果粒横径始终显著低于无核白。在整个果实发育期间,长粒无核白葡萄的果形指数始终显著高于无核白,花后16 d无核白葡萄的果形指数最大是1.80,而长粒无核白果实的果形指数在花后28 d达到最大,为2.12,果实发育前期果形指数已初步形成。图3~4
2.3.2 无核白和长粒无核白葡萄果实品质的差异
研究表明,长粒无核白葡萄的果穗质量、果穗长度、果穗宽度大于无核白,但与无核白葡萄间无显著差异。长粒无核白的果粒质量显著大于无核白葡萄,较无核白葡萄提高了48.94%。长粒无核白葡萄的可溶性固形物、VC含量高于无核白,但与无核白间无显著差异,长粒无核白(0.61 mg/100 g)的果实可滴定酸略低于无核白(0.64 mg/100 g),但无显著差异。长粒无核白葡萄的还原糖是283.57 mg/L,显著高于无核白葡萄(217.47 mg/L),长粒无核白葡萄还原糖高于无核白,可滴定酸低于无核白,导致其糖酸比(46.34)显著高于无核白葡萄(34.20)。表3
2.3.3 核白和长粒无核白葡萄果皮色差的差异
研究表明,长粒无核白和无核白葡萄L值、a值、b值、a/b、C值、h°、ΔE值间的差异不明显,均无显著差异。表4
2.4 无核白和长粒无核白葡萄干品质差异
2.4.1 无核白和长粒无核白葡萄干性状差异
研究表明,无核白的出干率和平均质量显著高于长粒无核白,出干率分别是26.41%、24.21%,平均质量分别是0.30、0.25 g。长粒无核白葡萄干果粒纵径显著高于无核白,横径显著低于无核白,致使长粒无核白的葡萄干果形指数显著高于无核白。表5
2.4.2 无核白和长粒无核白葡萄干品质的差异
研究表明,无核白葡萄干和长粒无核白葡萄干的品质差异较大。无核白葡萄干的可溶性固形物、还原糖、糖酸比均显著高于长粒无核白葡萄干,可滴定酸、维生素C含量显著低于对照。表6
2.4.3 无核白和长粒无核白葡萄干酚类物质差异
研究表明,长粒无核白葡萄干的总黄烷醇、总类黄酮、总单宁、总酚含量均显著高于无核白。长粒无核白葡萄干的总黄烷醇、总类黄酮、总酚含量是无核白葡萄干的2.68、2.62、1.77倍,长粒无核白总单宁比无核白增加了55.04%。表7
2.4.4 无核白和长粒无核白葡萄干抗氧化活性差异
研究表明,长粒无核白葡萄干的DPPH自由基清除能力(140.73 mg/kg)显著高于无核白(137.33 mg/kg);FRAP通过测定无核白葡萄干果实对Fe2+的还原能力评价其抗氧化能力,长粒无核白葡萄干对Fe2+的还原能力显著高于无核白,是无核白的1.59倍。无核白葡萄干ABTS自由基清除能力(257.63 mg/kg)略高于长粒无核白(256.17 mg/kg),但无显著差异。表8
2.5 葡萄干酚类物质和抗氧化活性相关性
研究表明,除ABTS外,总黄烷醇、总类黄酮、总单宁、总酚、DPPH、FRAP彼此之间的相关性较强,相关系数大于0.926,为显著或极显著差异。ABTS和总黄烷醇、总类黄酮、总单宁、总酚、DPPH、FRAP呈负相关,但无显著性差异。图5
3 讨 论
3.1 长粒无核白葡萄主要变异性状
芽变在葡萄中普遍存在,经过一系列的选择、培育可以育出新品种。红旗特早、瑞丰无核、户太系列、宇选1号均为优良芽变选育的新品种。芽变有整株芽变和局部芽变,经过2022~2023年系统观察发现,长粒无核白的芽变是局部芽变,主要表现在在嫩叶、嫩梢、叶柄颜色上。和普通无核白相比,长粒无核白幼叶正面颜色绿色带红条带,新梢腹侧颜色绿带红条带、节间背侧颜色绿带红条带、节间腹侧绿带红条带。
3.2 无核白和长粒无核白葡萄光合响应曲线差异
通过拟合分析,可以估算表观量子效率(a)、光补偿点(LCP)、光饱和点(LSP)、最大净光合速率,暗呼吸速率(Rd)等光合参数[22]。研究光响应曲线模型有很多,如二次多项式模型、直角函数模型、直角双曲线、非直角双曲线等,但是不能直接计算出饱和光强。叶子飘模型[23-25]可克服上述缺点,因此该光响应曲线的拟合选择该模型。
表观量子效率是衡量植物光合作用光能转化效率的主要指标之一[26],值越高,则代表植物转化利用光能的能力越强。无核白葡萄的表观量子效率是0.11%,长粒无核白的表观量子效率是0.02%,说明无核白转化利用光能的能力较强。
光补偿点(LCP)和光饱和点(LSP)分别代表植物利用弱光和强光的能力,LCP越低说明植物利用弱光能力越强,LSP越高,则说明植物利用强光能力越强[27]。无核白的光补偿点是10.53 μmol/(m2·s),长粒无核白是6.78 μmol/(m2·s),说明长粒无核白葡萄比无核白葡萄更耐弱光;无核白葡萄的光饱和点是1 085.58 μmol/(m2·s),长粒无核白是2 029.79 μmol/(m2·s),说明长粒无核白葡萄利用强光能力优于无核白葡萄。可初步推断,长粒无核白葡萄不管在弱光条件下还是强光条件下,利用光能的能力较无核白强。
植物光饱和点和补偿点不是固定的数值,且随外界条件的变化而变动,CO2浓度升高,光补偿点降低[28]。白云岗等[29]研究发现,无核白葡萄的光饱和点是1 178.362 μmol/(m2·s),光补偿点是67.487 μmol/(m2·s),均高于此次测定数据,可能是外界环境条件导致的。
3.3 无核白和长粒无核白葡萄品质差异及市场
唐美玲等[30]对烟台地区不同蛇龙珠营养系果实品质进行分析,表明营养系在可溶性固形物、酸、VC、单粒重、香气成分方面存在差异。经分析发现,长粒无核白葡萄鲜果的单粒重、还原糖、可滴定酸、VC和无核白葡萄之间存在差异。长粒无核白葡萄的果粒质量、还原糖、糖酸比显著高于无核白。长粒无核白葡萄的VC含量高于无核白,可滴定酸低于无核白,但均和无核白无显著差异。除了植物学性状(嫩叶、嫩梢、叶柄颜色)、果实品质发生变异外,最引起关注的是果形指数的差异。前期研究表明,未经植物生长调节剂处理的无核白鲜果和葡萄干果形指数分别是1.20、2.13,长粒无核白分别是1.51、3.10,长粒粒无核白的果形指数(鲜果、葡萄干)均显著高于无核白[11]。经过GA3处理后,无核白葡萄干单粒质量是0.79 g,长粒无核白是1.21 g,长粒无核白葡萄干果形指数是2.53,果粒形状为指形,无核白果形指数是1.43,果粒形状为短椭圆形,均存在显著差异[10]。近年长粒无核白葡萄干价格稳定30~45元/kg,可见长粒无核白葡萄的经济效益远远高于无核白。别尔麦提布·依明[9]研究认为,长粒无核白丰产性较强,可扩大栽培面积。
3.4 无核白和长粒无核白葡萄干的保健功能
酚类物质是重要的次生代谢物质[31-32]。酚类物质具有抗氧化、降血糖、抗增殖等多种生物活性功能[33]。Javanmardi等[34]研究发现,抗氧化活性和酚类含量呈正相关(R2=0.71)。Meng等[35]以新疆9个葡萄干为试材,开展了抗氧化活性、总酚及单体酚的研究,结果发现,Desert King葡萄的总酚、类类黄酮、总黄烷醇含量最高,DPPH抗氧化活性也显著高于其他品种。María等[36]研究了葡萄干晾制过程中酚类物质的变化,Hasim等[37]以晒干的葡萄干为试材,对酚类物质含量和抗氧化进了比较评价,结果发现不同品种间抗氧化活性差异较大,抗氧化活性的强弱和酚类物质含量关系较大。试验研究发现,长粒无核白葡萄干的总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力、对Fe2+的还原能力均显著高于葡萄无核白葡萄干。
卢登洋等[38]、蔡跃月等[39]、韩晓云等[40]的研究结果发现,总黄酮、总酚与DPPH自由基清除能力均存在极显著正相关。试验研究也发现,总黄烷醇、总类黄酮、总单宁、总酚、DPPH、FRAP彼此之间的相关系数大于0.926,为显著或极显著差异(P<0.05,P<0.01,P<0.001),与前人研究结果一致;试验研究还发现,ABTS和总黄烷醇、总类黄酮、总单宁、总酚、DPPH、FRAP呈负相关,但无显著性差异,与卢登洋等[38]、蔡跃月等[39]、韩晓云等[40]、李志荣等[41]的研究结果不一致。
4 结 论
4.1
和葡萄无核白葡萄相比,长粒无核白的变异主要表现在在嫩叶、嫩梢、叶柄颜色上。长粒无核白葡萄幼叶正面颜色绿色带红条带,新梢腹侧颜色绿带红条带、节间背侧颜色绿带红条带、节间腹侧绿带红条带。
4.2 长粒无核白葡萄不管在弱光条件下还是强光条件下,利用光能的能力强于无核白。
4.3 长粒无核白葡萄鲜果的单粒重、还原糖、可滴定酸、VC和无核白葡萄之间存在差异。果粒质量、还原糖、糖酸比均显著高于无核白。维生素C含量高于无核白,可滴定酸低于无核白,但均和无核白无显著差异。
4.4 长粒无核白葡萄干的总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力、FRAP(对Fe2+的还原能力)均显著高于无核白。
4.5 葡萄干总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力、FRAP(对Fe2+的还原能力)彼此之间的相关系数大于0.926,为显著或极显著差异,而ABTS和总黄烷醇、总类黄酮、总单宁、总酚、DPPH自由基清除能力、FRAP(对Fe2+的还原能力)呈负相关,但无显著差异。
参考文献(References)
[1]孔庆山. 中国葡萄志[M]. 北京: 中国农业科学技术出版社, 2004.
KONG Qingshan. Grapevine of China[M]. China Agricultural Science and Technology Press, 2004.
[2] 徐桂香, 廉苇佳, 刘萍, 等. 吐鲁番葡萄产业调研与分析[J]. 中外葡萄与葡萄酒, 2020,(3): 70-74.
XU Guixiang, LIAN Weijia, LIU Ping, et al. Investigation and analysis of the Turpan grape industry[J]. Sino-Overseas Grapevine amp; Wine, 2020,(3): 70-74.
[3] 王瑞华, 郭峰, 李海峰. 吐鲁番葡萄干产业发展分析及再升级策略[J]. 中外葡萄与葡萄酒, 2015,(6): 58-60.
WANG Ruihua, GUO Feng, LI Haifeng. Turpan raisin industry development analysis and re-upgrading strategy[J]. Sino-Overseas Grapevine amp; Wine, 2015,(6): 58-60.
[4] 卢诚. 浅谈葡萄的芽变选优[J]. 河北林业科技, 2009,(4): 92-93.
LU Cheng. Discussion on grape bud mutation selection[J]. The Journal of Hebei Forestry Science and Technology, 2009,(4): 92-93.
[5] 李志超, 赵诚. 长无核白葡萄的研究初报[J]. 果树科学, 1986, 3(2): 31-34, 49.
LI Zhichao, ZHAO Cheng. Preliminary report on the study of long-berry Thompson Seedless grape[J]. Journal of Fruit Science, 1986, 3(2): 31-34, 49.
[6] 李志超, 赵诚. 大无核白葡萄的研究初报[J]. 果树科学, 1986, 3(4): 36-41.
LI Zhichao, ZHAO Cheng. Preliminary report on the study of large seedless white grape[J]. Journal of Fruit Science, 1986, 3(4): 36-41.
[7] 闫鹏, 骆强伟, 廖康, 等. 无核白葡萄芽变系的植物学性状特征及RAPD标记研究[J]. 新疆农业科学, 2010, 47(1): 110-114.
YAN Peng, LUO Qiangwei, LIAO Kang, et al. Research on botanical characteristers and RAPD marker of Thomson seedless bud variation[J]. Xinjiang Agricultural Sciences, 2010, 47(1): 110-114.
[8] 张宗勤, 孙锋, 金强, 等. 欧洲葡萄品种无核白不同营养系果实的酚类物质[J]. 中国农业科学, 2011, 44(17): 3594-3602.
ZHANG Zongqin, SUN Feng, JIN Qiang, et al. Studies on the Phenolic Compounds in Berries of Vitis vinefera cv.Thompson Seedless Lines[J]. Scientia Agricultura Sinica, 2011, 44(17): 3594-3602.
[9] 别尔麦提布·依明, 廖康, 郝庆, 等. 无核白葡萄及其芽变品系的生长结实特性观察[J]. 新疆农业科学, 2013, 50(3): 440-446.
Bieermaitibu Yiming, LIAO Kang, HAO Qing, et al. Observation characteristics of growth and fruiting of white seedless and bud-sport strains[J]. Xinjiang Agricultural Sciences, 2013, 50(3): 440-446.
[10] 薛锋, 潘绪兵, 魏登攀, 等. 5个无核葡萄品种在吐鲁番地区的栽培性状及制干特性分析[J]. 新疆农业科学, 2023, 60(1): 105-115.
XUE Feng, PAN Xubing, WEI Dengpan, et al. Analysis on cultivation characteristics and raisin characteristics of five seedless grape cultivars in Turpan Region[J]. Xinjiang Agricultural Sciences, 2023, 60(1): 105-115.
[11] 户金鸽, 白世践. 无核白葡萄营养系果实品质及葡萄干特性分析[J]. 新疆农业科学, 2023, 60(11): 2751-2763.
HU Jinge, BAI Shijian. Study on the fruit quality and raisins character of Thompson seedless and Its lines[J]. Xinjiang Agricultural Sciences, 2023, 60(11): 2751-2763.
[12] NY/T 2563-2014.植物新品种特异性、一致性和稳定性测试指南 葡萄 [S].
NY/T 2563-2014.Guidelines for the conduct of tests for distinctness, uniformity and stability-grapevine (Vitis L.)[S].
[13] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006.
GAO Junfeng. Experimental guidance in plant physiology[M]. Beijing: Higher Education Press, 2006.
[14] Miller G L. Use of dinitrosalicylic acid reagent for determination of reducing sugar[J]. Analytical Chemistry, 1959, 31(3): 426-428.
[15] 谢辉, 闫鹏, 张雯, 等. 新疆无核白与无核白鸡心葡萄干品质特性研究[J]. 食品科学技术学报, 2016, 34(1): 37-41.
XIE Hui, YAN Peng, ZHANG Wen, et al. Study on quality characteristics of Xinjiang Thompson seedless and centennial seedless raisin[J]. Journal of Food Science and Technology, 2016, 34(1): 37-41.
[16] Mercurio M D, Dambergs R G, Herderich M J, et al. High throughput analysis of red wine and grape phenolics-adaptation and validation of methyl cellulose precipitable tannin assay and modified Somers color assay to a rapid 96 well plate format[J]. Journal of Agricultural and Food Chemistry, 2007, 55(12): 4651-4657.
[17] Tian S F, Wang Y, Du G, et al. Changes in contents and antioxidant activity of phenolic compounds during gibberellin-induced development in Vitis vinifera L. Muscat[J]. Acta Physiologiae Plantarum, 2011, 33(6): 2467-2475.
[18] Li Y G, Tanner G, Larkin P. The DMACA-HCl protocol and the threshold proanthocyanidin content for bloat safety in forage legumes[J]. Journal of the Science of Food and Agriculture, 1996, 70(1): 89-101.
[19] Wolfe K, Wu X Z, Liu R H. Antioxidant activity of apple peels[J]. Journal of Agricultural and Food Chemistry, 2003, 51(3): 609-614.
[20] 张妍, 孙娟, 包菲菲, 等. 大兴安岭地区蓝果忍冬果实多酚鉴定及生物活性分析[J]. 食品科学, 2023, 44(12): 225-234.
ZHANG Yan, SUN Juan, BAO Feifei, et al. Identification and bioactivity analysis of polyphenols in blue honeysuckle fruit from different regions of the greater khingan mountains[J]. Food Science, 2023, 44(12): 225-234.
[21] Benzie I F F, Strain J J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay[J]. Analytical Biochemistry, 1996, 239(1): 70-76.
[22] 周晓旭, 张玉翠, 吴林, 等. 冬小麦光合特性对CO2浓度与土壤含水量的响应机制[J]. 中国生态农业学报, 2017, 25(7): 1034-1041.
ZHOU Xiaoxu, ZHANG Yucui, WU Lin, et al. Response mechanism of photosynthetic characteristics of winter wheat to CO2 concentration and soil water content[J]. Chinese Journal of Eco-Agriculture, 2017, 25(7): 1034-1041.
[23] 叶子飘, 于强. 一个光合作用光响应新模型与传统模型的比较[J]. 沈阳农业大学学报, 2007, 38(6): 771-775.
YE Zipiao, YU Qiang. Comparison of A new model of light response of photosynthesis with traditional models[J]. Journal of Shenyang Agricultural University, 2007, 38(6): 771-775.
[24] 叶子飘, 于强. 光合作用光响应模型的比较[J]. 植物生态学报, 2008, 32(6): 1356-1361.
YE Zipiao, YU Qiang. Comparison of new and several classical models of photo-synthesis in response to irradiance[J]. Journal of Plant Ecology, 2008, 32(6): 1356-1361.
[25]叶子飘, 李进省. 光合作用对光响应的直角双曲线修正模型和非直角双曲线模型的对比研究[J]. 井冈山大学学报(自然科学版), 2010, 31(3): 38-44.
YE Zipiao, LI Jinsheng. Comparative investigation light response of photosynthesis on non-rectangular Hyperbola model and modified model of rectangular Hyperbola[J]. Journal of Jinggangshan University (Natural Science), 2010, 31(3): 38-44.
[26] Yu Q, Zhang Y Q, Liu Y F, et al. Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2 changes[J]. Annals of Botany, 2004, 93(4): 435-441.
[27] Long S P, Humphries S, Falkowski P G. Photoinhibition of photosynthesis in nature[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1994, 45: 633-662.
[28] 王忠. 植物生理学[J].北京中国农业出版社2008.
WANG Zhong. Plant physiology[M]. Beijing: China Agriculture Press, 2008.
[29] 白云岗, 刘洪波, 张江辉, 等. 葡萄光合作用光响应曲线拟合模型比较研究[J]. 节水灌溉, 2016,(9): 8-11.
BAI Yungang, LIU Hongbo, ZHANG Jianghui, et al. Comparison of light-response curve fitted models for grape[J]. Water Saving Irrigation, 2016,(9): 8-11.
[30] 唐美玲, 宋来庆, 张超杰, 等. 烟台地区不同蛇龙珠营养系果实品质分析[J]. 中外葡萄与葡萄酒, 2011,(7): 9-13.
TANG Meiling, SONG Laiqing, ZHANG Chaojie, et al. Analysis of the fruit quality of different Cabernet Gernischt clones in Yantai district[J]. Sino-Overseas Grapevine amp; Wine, 2011,(7): 9-13.
[31] Guo R X, Guo X B, Li T, et al. Comparative assessment of phytochemical profiles, antioxidant and antiproliferative activities of Sea buckthorn (Hippopha? rhamnoides L.) berries[J]. Food Chemistry, 2017, 221: 997-1003.
[32] Swallah M S, Sun H, Affoh R, et al. Antioxidant potential overviews of secondary metabolites (polyphenols) in fruits[J]. International Journal of Food Science, 2020, 2020: 9081686.
[33] Sharma A, Kim J W, Ku S K, et al. Anti-diabetic effects of blue honeyberry on high-fed-diet-induced type II diabetic mouse[J]. Nutrition Research and Practice, 2019, 13(5): 367-376.
[34] Javanmardi J, Stushnoff C, Locke E, et al. Antioxidant activity and total phenolic content of Iranian Ocimum accessions[J]. Food Chemistry, 2003, 83(4): 547-550.
[35] Meng J F, Fang Y L, Zhang A, et al. Phenolic content and antioxidant capacity of Chinese raisins produced in Xinjiang Province[J]. Food Research International, 2011, 44(9): 2830-2836.
[36] Fabani M P, Baroni M V, Luna L, et al. Changes in the phenolic profile of Argentinean fresh grapes during production of Sun-dried raisins[J]. Journal of Food Composition and Analysis, 2017, 58: 23-32.
[37] Kelebek H, Jourdes M, Selli S, et al. Comparative evaluation of the phenolic content and antioxidant capacity of Sun-dried raisins[J]. Journal of the Science of Food and Agriculture, 2013, 93(12): 2963-2972.
[38] 卢登洋, 王鑫, 唐章虎, 等. 梨果实发育过程中酚类物质组成及抗氧化活性比较[J]. 中国农业科技导报, 2023, 25(9): 97-104.
LU Dengyang, WANG Xin, TANG Zhanghu, et al. Phenolic composition and antioxidant activity of pear species during fruit development comparison[J]. Journal of Agricultural Science and Technology, 2023, 25(9): 97-104.
[39] 蔡跃月, 麦尔哈巴·阿布拉, 高路, 等. 滇红玫瑰发酵过程中酚类物质含量及其抗氧化和抗炎活性分析[J]. 食品工业科技, 2024, 45(11): 213-221.
CAI Yueyue, Maierhaba Abula, GAO Lu, et al. Analysis of phenolic content and its antioxidant and anti-inflammatory activities during the fermentation process of Rosa rugosa Dianhong[J]. Science and Technology of Food Industry, 2024, 45(11): 213-221.
[40] 韩晓云, 陶雨婷, 战佳莹, 等. 桑葚发酵前后酚类组成变化及其抗氧化活性分析[J]. 食品工业科技, 2024, 45(2): 280-288.
HAN Xiaoyun, TAO Yuting, ZHAN Jiaying, et al. Analysis of phenolic composition changes and antioxidant activity of mulberry before and after fermentation[J]. Science and Technology of Food Industry, 2024, 45(2): 280-288.
[41] 李志荣, 邵起菊, 吴其妹, 等. 不同成熟度老鹰茶中酚类化合物含量及抗氧化活性研究[J]. 广西植物, 2024, 44(6): 1170-1181.
LI Zhirong, SHAO Qiju, WU Qimei, et al. Study on the Content and Antioxidant Activity of Phenolic Compounds in Eagle Tea with Different Maturity [J].Guangxi Botany, 2024, 44 (6): 1170-1181.
Difference analysis between thompson seedless and bud mutation long-berry thompson seedless grape
HU Jinge1, BAI Shijian1, ZHENG Ming2
(1. Research Institute of Grapes and Melons of Xinjiang Uygur Autonomous Region, Shanshan Xinjiang 838200,China; 2. Xinjiang Research Institute of Water Resources and Hydropower, Urumqi 830049, China)
Abstract:【Objective】 To analyze the differences of botanical characters, berry quality, raisin polyphenols and antioxidant activity between Thompson Seedless and long-berry Thompson Seedless grape in the hope of providing new germplasm resources for grape breeding.
【Methods】" V.vinefera cv.Thompson Seedless and bud mutation Long-berry Thompson Seedless were used as material, in the open field of the experimental vineyard,to investigate botanical character, photosynthetic response curve, berry quality and raisins quality and analyze the relationship between phenolic and antioxidant activity of raisins.
【Results】" The results showed that, the main variation characteristics of bud mutation Long-berry Thompson Seedless were the color of young leaves, tender tips and petioles.Compared with ordinary Thompson Seedless, the color of young leaves was green with red bands, ventral new shoot color was green with red bands, dorsal internode color was green with red bands, and intersegmental ventral was green with red band.Germination rate of Thompson Seedless was higher than Long-berry Thompson Seedless, which were 65.00% and 63.82% respectively, germination rates of Long-berry Thompson Seedless were 50.48% and 53.45% respectively, in 2022, in 2023.The shape index of Long-berry Thompson Seedless were always higher than that of Thompson Seedless, the maximum shape index of Thompson Seedless and Long-berry Thompson Seedless were 1.80 and 2.12, after 16 d and 28 d after flowing, respectively.Berry quality, reduce sugar, sugar-acid ratio of Thompson Seedless were significantly higher than those of Long-berry Thompson Seedless, and there were no difference for bunch weight, bunch length, bunch width, soluble solid, titratable acid, Vitamin C, chromatic aberration.Drying rate, mean weight of Thompson Seedless were significantly higher than those of Long-berry Thompson Seedless, shape index of Long-berry Thompson Seedless was significantly higher than that of Thompson Seedless.Total flavanol, total flavonoids, total tannin, total phenol of Long-berry Thompson Seedless were significantly higher than those of Thompson Seedless, and their content were 1,808.59、2,286.43, 51.11, 98.53 mg/kg, respectively.DPPH and FRAP free radical scavenging capacity of Long-berry Thompson Seedless were significantly higher than Thompson Seedless, ABTS free radical scavenging capacity of Long-berry Thompson Seedless lower than Thompson Seedless, and no difference.The correlation coefficient were greater than 0.926 for total flavanol, total flavonoids, total tannin, total phenol, DPPH and FRAP free radical scavenging capacity, and their differences were significant or extremely significant.ABTS free radical scavenging capacity was negatively correlated with total flavanol, total flavonoids, total tannins, total phenols, DPPH and FRAP free radical scavenging capacity with litter difference.
【Conclusion】" Bud mutation Long-berry Thompson Seedless can be used as an excellent variety and provide new germplasm resources for breeding.
Key words:thompson seedless; long-berry thompson seedless; fruit quality; raisin
Fund projects:Project of Natural Science Foundation of Xinjiang Uygur Autonomous Region (2022D01A120); Major Scientific R amp; D Program Project of Xinjiang Uygur Autonomous Region(2022B02045-2); Basic Scientific R amp; D Program of Public Welfare Research Institutes of Xinjiang Uygur Autonomous Region of China (KY2023099); Xinjiang Uygur Autonomous Region \"Tianshan Talent\" Training Program \"Agriculture, Rural Areas, and Farmers\" Backbone Talent(2023SNGGNT062)
Correspondence author: BAI Shijian (1986-), male, from Yunnan, senior agronomist with undergraduate degree, research direction: grape cultivation, (E-mail) 594748964@qq.com
基金项目:新疆维吾尔自治区自然科学基金项目(2022D01A120);新疆维吾尔自治区重点研发计划专项(2022B02045-2);新疆维吾尔自治区公益性科研院所基本科研业务费专项资金(KY2023099);新疆维吾尔自治区“天山英才”培养计划“三农”骨干人才项目(2023SNGGNT062)
作者简介:户金鸽(1982-),女,新疆塔城人,副研究员,硕士,研究方向为葡萄育种与栽培,(E-mail)hujinge2007@sina.com
通讯作者:白世践(1986-),男,云南人,高级农艺师,研究方向为葡萄育种与栽培,(E-mail)594748964@qq.com